Биохимия и молекулярная биология - Белясова Н.А. 2002
Метаболизм. Процессы, требующие притока энергии
Особенности биосинтеза. Биосинтез углеводов
Закономерности обмена и биосинтеза моносахаридов
Ключевым промежуточным соединением в обмене углеводов можно назвать глюкозо-6-фосфат. Это вещество может образовываться из экзогенной глюкозы при ее фосфорилировании в процессе транспорта через мембраны или сразу после этого. Кроме того, глюкозо-6-фосфат образуется в клетках автотрофных организмов, способных к фиксации СО2, в процессе глюконеогенеза из глицеральдегид-3-фосфата (рис. 13.5). Другими источниками глюкозо-6-фосфата являются: пируват (образуется в результате большинства катаболитных путей расщепления углеводов), некоторые промежуточные продукты ЦТК, углеродные скелеты гликогенных аминокислот, лактат (продукт молочнокислого брожения), глицерол (образуется при расщеплении липидов). Все эти вещества могут вовлекаться в процесс глюконеогенеза — синтеза глюкозы из неуглеводных предшественников. Кроме этого, в клетках растений и многих микроорганизмов глюкоза может образовываться из ацетил-СоА — продукта расщепления жирных кислот. В этом случае используются два специфических фермента глиоксилатного цикла, отсутствующие в клетках высших животных. Эти ферменты (изоцитрат-лиаза и малат-синтаза) участвуют в превращении ацетил-СоА в сукцинат, который затем преобразуется в оксалоацетат (рис. 11.7) и вступает в процесс глюконеогенеза. Наконец, еще одним источником глюкозы, а значит, и глюкозо-6-фосфата являются резервные полисахариды.
Расходуется глюкозо-6-фосфат в клетках следующим образом. Во-первых, он является субстратом разнообразных катаболических путей, в которых запасается энергия, образуются восстановительные эквиваленты и строительные блоки (глава 9). Во-вторых, глюкозо-6-фосфат способен изомеризоваться в глюкозо-1-фосфат и служить субстратом для биосинтеза гликогена либо иных запасных и структурных полисахаридов. Глюкозо-6-фосфат может дефосфорилироваться с образованием глюкозы, которая способна трансформироваться в клетках в другие моносахариды, например галактозу, маннозу, фруктозу, глюкуроновую кислоту. Названные соединения принимают участие в процессах образования сложных полисахаридов клеточных стенок или других структур.
Большинство перечисленных выше способов образования и расходования моносахаридов уже охарактеризовано в предыдущих разделах. Здесь внимание будет уделено одному из центральных путей образования ключевого продукта углеводного обмена (глюкозо-6-фосфата) — глюконеогенезу.
Глюконеогенез. Этот процесс характерен для представителей всех царств живых организмов, но наиболее важное значение имеет для клеток высших животных. Дело в том, что эмбриональные ткани, мозг, семенники, эритроциты в качестве источника углерода способны использовать только D-глюкозу. Если в рационе недостает углеводов, в печени индуцируется распад гликогена, но и этого источника может оказаться недостаточно (мозг человека в сутки потребляет более 120 г глюкозы). В таком случае глюкоза синтезируется в организме из неуглеводных предшественников в ходе глюконеогенеза. Наиболее активно глюконеогенез осуществляется у животных в клетках печени и почек.
Реакции глюконеогенеза в большой степени тождественны обратным реакциям гликолиза, и многие из них катализируются теми же ферментами, которые задействованы в гликолизе (рис 14.1).
Итак, в гликолизе имеется три практически необратимые реакции, взамен которых в глюконеогенезе существуют обходные пути. Первый обходной путь представляет собой превращение пирувата в фосфоенол-пируват. Для непосредственного перевода пирувата в фосфоенолпируват недостаточно энергии расщепления АТР, поэтому данная стадия осуществляется в ходе нескольких реакций. Вначале пируват, образующийся преимущественно в цитоплазме (из лактата, аминокислот, в гликолизе), переводится в митохондрии и там карбоксилируется в оксалоацетат (рис. 14.2). Катализирует реакцию пируваткарбоксилаза, использующая в качестве кофактора биотин. Эта анаплеротическая реакция и характеристика свойств фермента описаны в разделе 11.2.
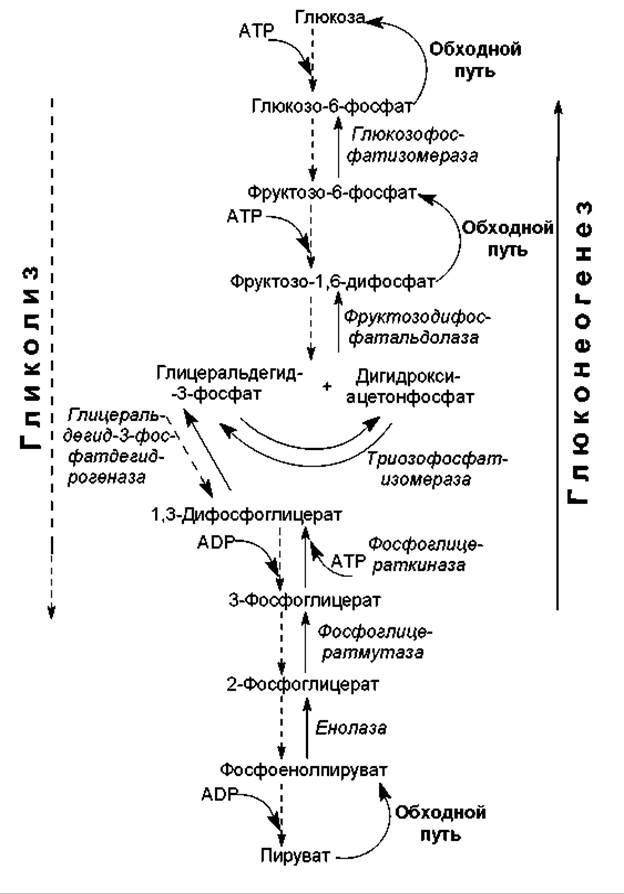
Рис. 14.1. Последовательность реакций гликолиза (пунктирные стрелки) и глюконеогенеза (сплошные стрелки). Подписаны названия ферментов, общих для двух путей. Стадии, по которым процессы различаются, обозначены в глюконеогенезе, как обходные пути
Оксалоацетат в митохондриях восстанавливается в малат (митохондриальная малатдегидрогеназа), который с помощью специфических переносчиков транспортируется в цитоплазму. В цитоплазме малат вновь окисляется в оксалоацетат (цитоплазматическая малатдегидрогеназа), который с помощью GTP-зависимой фосфоенолпируваткарбоксилазы декарбоксилируется в фосфоенолпируват (РЕР).
Второй обходной путь в глюконеогенезе представляет собой превращение фруктозодифосфата во вруктозо-6-фосфат (рис. 14.2). В гликолизе фосфофруктокиназная реакция, сопровождающаяся гидролизом АТР, является необратимой. В глюконеогенезе функционирует другой фермент — фруктозодифосфатаза, которая катализирует практически необратимое отщепление фосфатной группы от первого атома углерода. Фруктозодифосфатаза, как и пируваткарбоксилаза, является аллостерическим ферментом. Его активность ингибируется с помощью АМР и активируется при участии АТР.
Третий обходной путь — дефосфорилирование глюкозо-6-фосфата, не может произойти с помощью прямого обращения гексокиназной реакции. Эту реакцию катализирует глюкозо-6-фосфатаза, которая локализована на внутренней поверхности мембран гладкого эндоплазматического ретикулума (ЭР). Поэтому для осуществления данной реакции глюкозо-6-фосфат транспортируется в ЭР, где дефосфорилируется в свободную глюкозу (рис. 14.2). Следует отметить, что глюкозо-6-фосфатаза отсутствует в таких тканях, как мышцы и мозг, поэтому они не могут поставлять в кровь свободную глюкозу.
Суммарное уравнение глюконеогенеза выглядит следующим образом:
2Пируват + 4АТР + 2GTP +2NADH + 2H+ + 4H2O →
→ Глюкоза + 2NAD+ + 4ADP + 2GDP + 6 Pi
Из приведенного баланса следует, что на образование одной молекулы глюкозы в процессе глюконеогенеза расходуется шесть высокоэнергетических фосфатных связей, а также две молекулы NADH. Важно отметить, что регуляция скорости синтеза глюкозы в этом пути осуществляется с помощью ферментов, не принимающих участие в гликолизе. При этом глюконеогенез наиболее интенсивно протекает в условиях повышенного содержания в клетке топливных молекул, в частности ацетил-СоА, и достаточного количества АТР.
Глицерол включается в путь глюконеогенеза через дигидроксиацетонфосфат, в который он превращается после фосфорилирования (с участием АТР) и дегидрирования.
Аминокислоты поступают в путь через такие метаболиты, как пируват и оксалоацетат, образующиеся в ходе перестроек их углеродных скелетов. Лактат перед вступлением в глюконеогенез должен окислиться до пирувата.
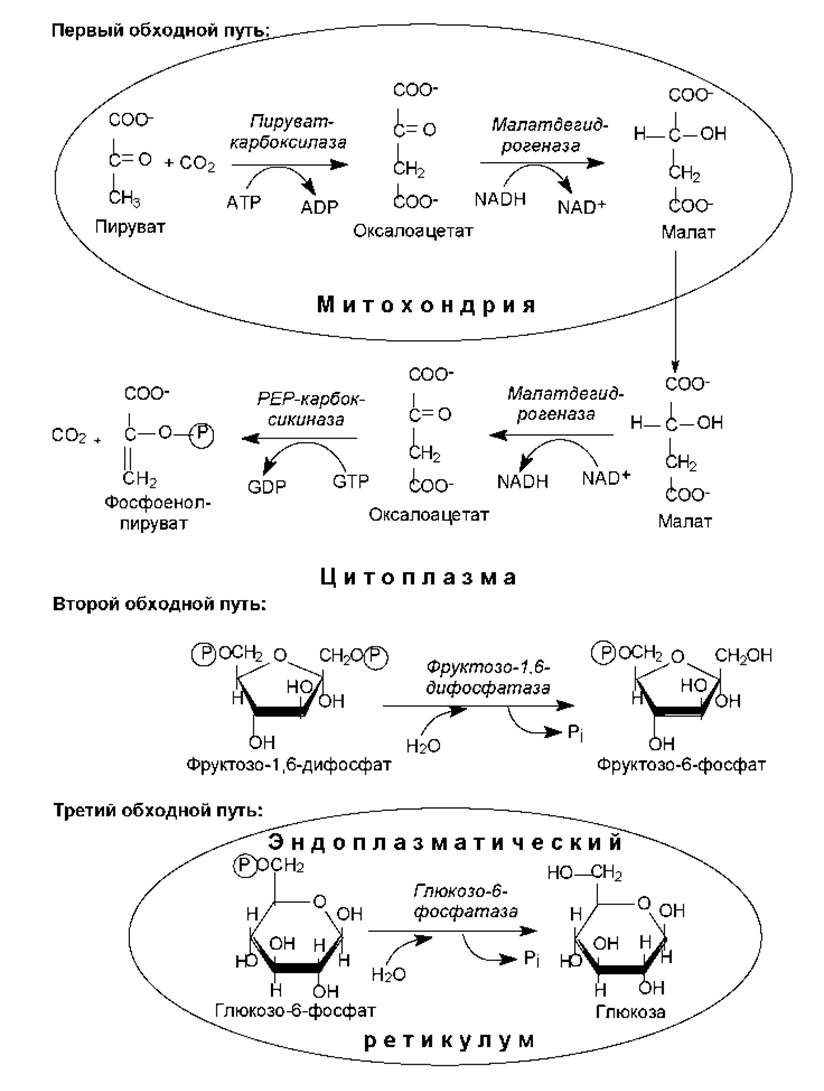
Рис. 14.2. Реакции обходных путей в глюконеогенезе