Биохимия и молекулярная биология - Белясова Н.А. 2002
Метаболизм. Процессы, требующие притока энергии
Биосинтез липидов
Биосинтез полярных и неполярных липидов
Биосинтез липидов в клетках эукариот осуществляется на мембранах гладкого эндоплазматического ретикулума. Большинство ферментов, принимающих участие в этих процессах, ассоциированы с его мембранами и представляют собой липопротеины.
Основными предшественниками для синтеза нейтральных липидов (триацилглицеролов), а также полярных фосфо- и гликолипидов служат активированные жирные кислоты и глицерол-3-фосфат. Активация жирных кислот происходит в следующей реакции:
Жирная кислота + АТР + СоА → Ацил-S-СоА + АМР + PPi
Глицерол-3-фосфат образуется либо при прямом фосфорилировании за счет АТР при участии глицеролкиназы, либо при восстановлении промежуточного продукта гликолиза — дигидроксиацетон-3-фосфата ферментом 3-глицерофосфатдегидрогеназой, использующей в качестве кофермента NADH.
Биосинтез неполярных липидов. Если биосинтез триацилглицеролов осуществляется de novo (из глицеролфосфата и жирных кислот), то на первой стадии происходит последовательное ацилирование двух свободных гидроксильных групп молекулы глицерол-3-фосфата (рис. 15.3). При этом в реакции этерификации СоА-эфиром жирной кислоты по первому атому углерода глицерол-3-фосфата вначале формируется моноацилглицерол-3-фосфат (лизофосфатид), а затем — диацилглицерол-3-фосфат (фосфатидат).
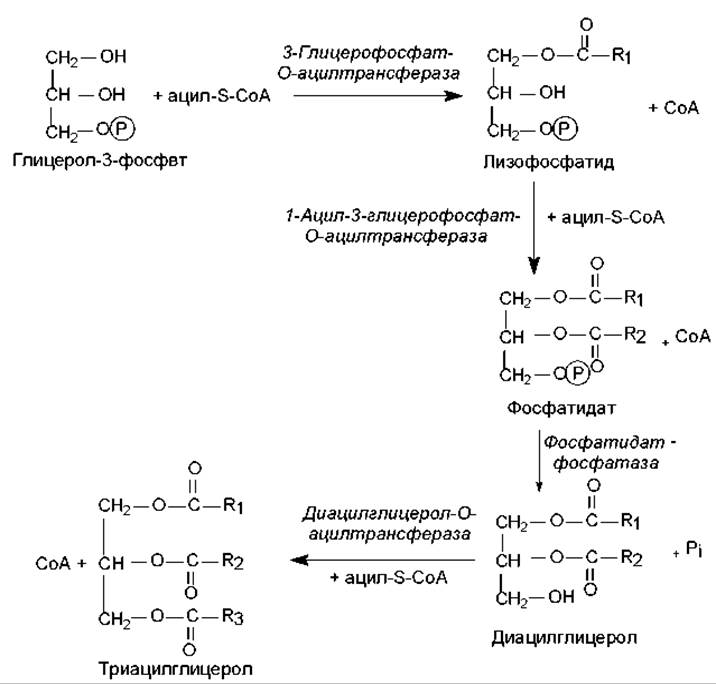
Рис. 15.3. Биосинтез триацилглицеролов: R1, R2, R3 — углеводородные цепи жирных кислот
На следующей стадии происходит гидролитическое отщепление фосфатной группы от молекулы фосфатидата и образуется 1, 2-диацилглицерол, который взаимодействует с третьим СоА-производным жирной кислоты, в результате чего формируется триацилглицерол (триглицерид).
Кроме описанной схемы, синтез нейтральных липидов может осуществляться с участием в качестве предшественников продуктов расщепления липидов, попадающих в организм с пищей. Эти процессы особенно интенсивны в слизистой кишечника животных. Нейтральные жиры расщепляются в пищеварительном тракте панкреатическими липазами до жирных кислот и 2-моноацилглицеролов, которые всасываются слизистой кишечника. В клетках слизистой оболочки происходит последовательное ацилирование 2- моноацилглицерола СоА-эфирами жирных кислот с образованием триацилглицеролов. Эти реакции катализируют особые ацилтрансферазы.
Триглицериды, как уже отмечалось, являются основными запасными веществами в клетках животных и некоторых других организмов. Особое значение они имеют для впадающих в спячку и мигрирующих на далекие расстояния животных. Например, верблюды запасают триацилглицеролы в горбу и используют их как источник воды, которая образуется при окислении. У полярных животных (тюленей, моржей и др.) триглицериды часто выполняют функцию теплоизолятора. Некоторые животные используют неполярные липиды для регулирования плавучести. Например, в спермацетовом мешке кашалотов находится несколько тонн триацилглицеролов, содержащих в составе молекул ненасыщенные жирные кислоты. Плотность (консистенция) этих триглицеридов зависит от температуры среды: повышается при понижении температуры. Питаясь кальмарами, кашалоты заплывают на большие глубины, где температура воды ниже обычной. Это индуцирует кристаллизацию триглицеридов, увеличивается их плотность соответственно увеличению плотности морской воды на глубине, и животное, не прибегая к дополнительным усилиям, может долго оставаться на большой глубине.
Биосинтез полярных липидов. Первые этапы биосинтеза фосфо- и гликолипидов совпадают с таковыми для синтеза триацилглицеролов: в ходе этих реакций тоже образуются фосфатидат и диацилглицерол (рис. 15.3). На следующих этапах к молекуле диацилглицерола может присоединяться с помощью специфического переносчика активированная полярная «голова» молекулы (чаще аминоспирт). В других случаях, наоборот, на полярную «голову» переносится активированная молекула диацилглицерола.
На рис. 15.4 представлены реакции биосинтеза фосфатидилхолина. В этом процессе активация холина осуществляется путем соединения с СDP, и этот нуклеозиддифосфат служит переносчиком холинфосфата на молекулу диацилглицерола. Подобная закономерность наблюдается в биосинтезе полисахаридов, только там переносится сам моносахарид, а не его фосфорилированная форма (рис. 14.3).
Аналогичным путем (с использованием цитидиндифосфатэтаноламина) синтезируется другой важный компонент мембран — фосфатидилэтаноламин.
Другие фосфолипиды, такие, как фосфатидилинозит, фосфатидилглицерол, дифосфатидилглицерол, фосфатидилсерин, синтезируются при участии активированного диацилглицерола — цитидиндифосфатдиацилглицерола. Это соединение образуется при взаимодействии СТР с фосфатидатом (рис. 15.5). Перенос остатка фосфодиацилглицерола на один из спиртов катализируется специфическими фосфатидилтрансферазами и приводит к формированию перечисленных выше глицерофосфолипидов.
Кроме охарактеризованных выше способов биосинтеза полярных липидов, существует возможность взаимопревращения глицерофосфолипидов между собой (рис. 15.5). Так при декарбоксилировании фосфатидилсерина образуется фосфатидилэтаноламин, а он, в свою очередь, может превращаться в фосфатидилхолин при троекратном метилировании S-аденозилметионином атома азота. Фосфатидилглицерол-1-фосфат способен превращаться в дифосфатидилглицерол.
Описанные закономерности биосинтеза глицерофосфолипидов характерны и для процессов синтеза других полярных липидов. На скорость биосинтеза липидов у животных очень сильно воздействуют гормоны, в первую очередь инсулин, который стимулирует синтез жирных кислот из глюкозы (продуктов ее расщепления).
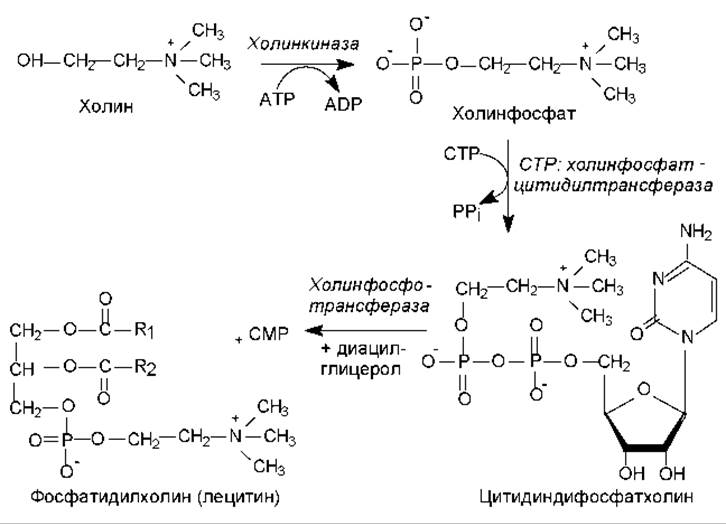
Рис. 15.4. Биосинтез фосфатидилхолина: R1, R2 — углеводородные цепи жирных кислот; СТР — цитидинтрифосфат
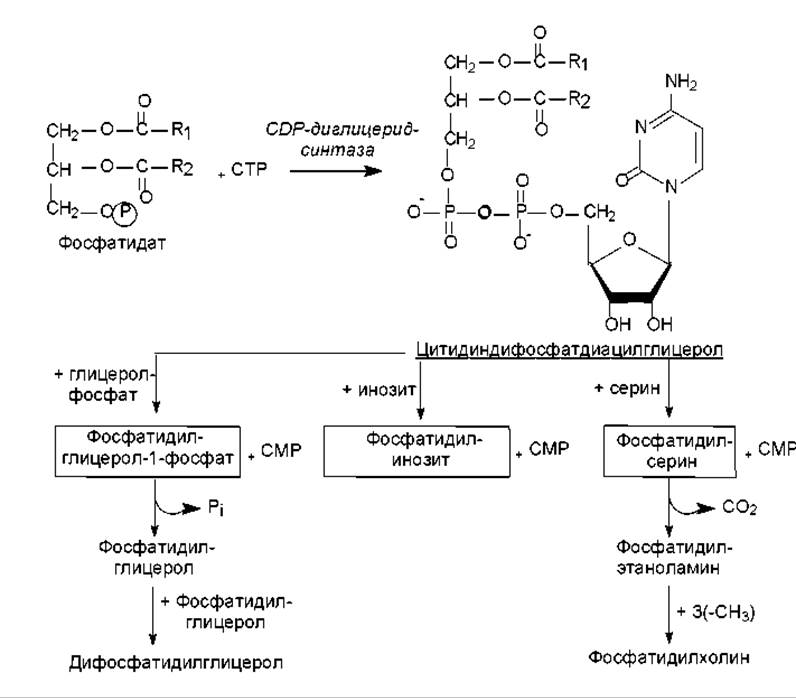
Рис. 15.5. Альтернативные пути биосинтеза полярных липидов
Биосинтез стеролов. Эти соединения могут синтезировать все организмы, однако многие из них используют стеролы, присутствующие в пище. Например, суточная потребность человека в холестероле удовлетворяется на 50% с продуктами питания, и на 50% — за счет биосинтеза.
Основным предшественником для биосинтеза стеролов является ацетил-СоА, который поэтапно, с соблюдением определенных стадий (правило Ружечки) превращается в фарнезилпирофосфат (рис. 17.2), служащий также промежуточным соединением в синтезе каротиноидов. При димеризации двух молекул фарнезилпирофосфата по типу «голова к голове» возникает 30-углеродный линейный изопреноид сквален, который циклизуется и модифицируется с образованием стеролов (рис. 15.6). Данный процесс идентичен синтезу каротиноидов, и его принципы, а также этапы биосинтеза фарнезилпирофосфата, описаны в главе 17.
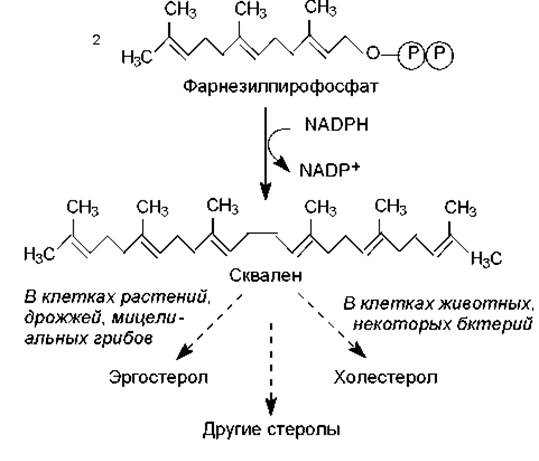
Рис. 15.6. Некоторые этапы биосинтеза стеролов
Эргостерол получают биотехнологическим путем при культивировании дрожжей (Saccharomyces cerevisiae, Saccharomyces carlbergensis, Candida guilliermondii), а также мицелиальных грибов (Penicillium notatum). Эргостерол является предшественником эргокальциферола (витамина D2), а холестерол, синтезируемый животными и некоторыми бактериями, служит предшественником холекальциферола (витамина D3).