БИОХИМИЯ - В. В. Емельянов - 2016
РАЗДЕЛ 5. ОБМЕН БЕЛКОВ И НУКЛЕИНОВЫХ КИСЛОТ
5.1. Биологическая роль белков
В количественном отношении белки образуют самую важную группу макромолекул. Белкам присущ ряд уникальных функций:
- биокаталитическая, все ферменты - вещества белковой природы;
- пластическая, белки входят в состав важнейших структур клеток и тканей;
- регуляторная, большинство гормонов имеют белково-пептидную природу;
- защитная, белки - иммуноглобулины - обеспечивают защиту организма от действия чужеродных антигенов;
- дыхательная, белок гемоглобин в составе эритроцитов обеспечивает транспорт кислорода в ткани и СО2 в легкие;
- транспортная, перенос многих важных для организма веществ по крови происходит в комплексе с белками.
В организме человека массой 70 кг содержится примерно 10 кг белка, суточная норма белков в питании составляет примерно 100 г. Доля других азотсодержащих соединений в организме невелика, поэтому и баланс азота в организме определяется прежде всего метаболизмом белков. В отличие от углеводов и липидов белки и составляющие их аминокислоты не способны резервироваться в организме.
5.2. Превращения белков и аминокислот в организме
Белки, поступившие в организм с пищей, в желудочно-кишечном тракте расщепляются до аминокислот под действием протеолитических ферментов (пептид- гидролазы, пептидазы, протеазы). Внутренние пептидные связи расщепляются эндопептидазами (пепсин, трипсин и химотрипсин), концевые - экзопептидазами (карбоксипептидазы и аминопептидазы). В совокупности эндо- и экзопептидазы доводят гидролиз белков до образования аминокислот.
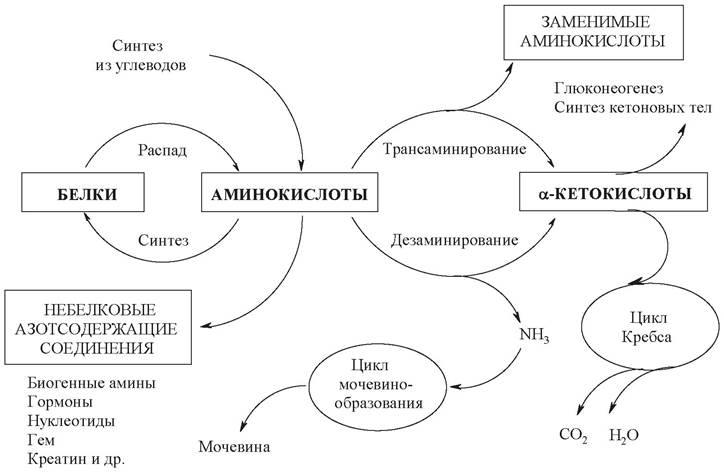
Образовавшиеся аминокислоты затем всасываются стенкой кишечника и после транспорта кровью включаются в клетках в различные пути использования, главным из которых является синтез собственных белков. Значительная часть аминокислот разносится кровью по всему организму и используется для физиологических целей. Аминокислоты также участвуют в синтезе специфических азотсодержащих соединений - пуриновых и пиримидиновых нуклеотидов, креатина и др. Азот аминокислот, отщепляемый на ранних стадиях катаболизма, в зависимости от потребности организма может утилизироваться в анаболических процессах, а также включаться в конечный продукт обмена азота - мочевину и экскретироваться из организма. Безазотистые углеродные остатки аминокислот образуют чаще всего кетокислоты, которые далее деградируют по общим путям катаболизма других окисленных углеводородов. Включаясь в общие пути катаболизма, аминокислоты могут служить источником энергии. Основные пути использования аминокислот представлены на рис. 21.
Рис. 21. Основные пути использования аминокислот
В организме создается фонд аминокислот, который пополняется из следующих источников:
- аминокислоты пищи;
- аминокислоты, образующиеся при катаболизме собственных белков;
- аминокислоты, синтезируемые самими клетками.
При этом возможности синтеза аминокислот в организме существенно ограничены. В табл. 7 указаны незаменимые аминокислоты, которые не могут быть синтезированы клетками из-за отсутствия источников синтеза, поэтому их присутствие в белках пищи обязательно. Выделяют также частично заменимые аминокислоты, которые могут синтезироваться в небольших количествах в организме, и условно заменимые аминокислоты, для синтеза которых необходимы незаменимые аминокислоты. Потребность в заменимых аминокислотах может быть восполнена синтезом из других соединений.
Таблица 7. Заменимые и незаменимые аминокислоты
Незаменимые |
Условно заменимые |
Частично заменимые |
Заменимые |
Валин |
Тирозин |
Аргинин |
Аланин |
Изолейцин |
Цистеин |
Гистидин |
Аспарагин |
Лейцин |
Аспартат |
||
Лизин |
Глицин |
||
Метионин |
Глутамат |
||
Треонин |
Глутамин |
||
Триптофан |
Пролин |
||
Фенилаланин |
Серин |
Аминокислотный состав определяет биологическую ценность белка. Чем выше содержание незаменимых аминокислот, тем больше его пищевая ценность. Полноценными белками в питании считаются те, которые содержат незаменимые аминокислоты в нужном количественном и качественном соотношении.
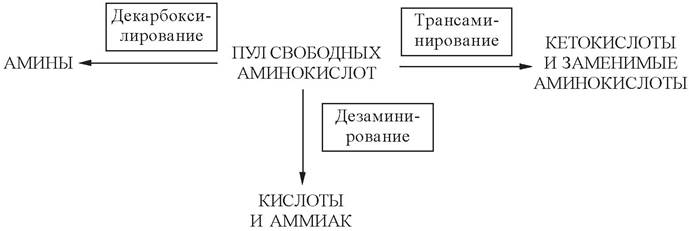
5.3. Общие пути обмена аминокислот
Общие пути обмена аминокислот включают реакции дезаминирования, транс- аминирования и декарбоксилирования:
Реакции трансаминирования (переаминирования) являются наиболее важными реакциями среди общих путей обмена аминокислот. Они играют основную роль в процессе глюконеогенеза и образования новых аминокислот. Суть этой реакции - обратимый перенос аминогруппы с аминокислоты на кетокислоту без промежуточного образования аммиака. Катализируют реакции трансаминированияферменты трансаминазы (аминотрансферазы), в роли кофермента выступает пиридоксальфосфат (активная форма витамина В6):
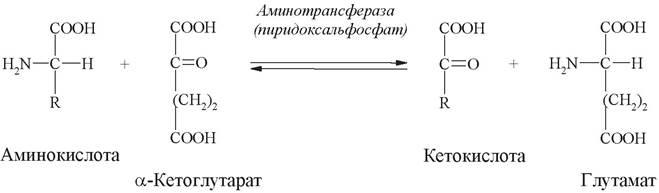
В переаминировании участвует много аминокислот (кроме лизина, треонина), наиболее активными являются глутаминовая, аспарагиновая кислоты, что связывают с высоким содержанием в тканях животных двух специфических аминотрансфераз - аспартатаминотрансферазы и аланинаминотрансферазы.
Аминотрансферазы участвуют в синтезе новых аминокислот из α-кетоглутарата. Этот синтез возможен и для получения незаменимых аминокислот, если с пищей будут поступать соответствующие кетокислоты.
Дезаминирование - реакция, в ходе которой аминогруппа освобождается в виде аммиака. Механизмы дезаминирования могут быть различными:
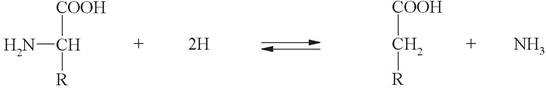
а) восстановительное дезаминирование (с образованием насыщенной жирной кислоты):
б) гидролитическое дезаминирование (с образованием карбоновой гидроксикислоты):

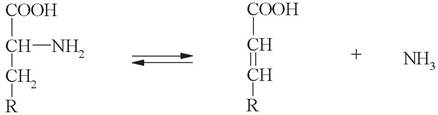
в) элиминирующее дезаминирование (с образованием ненасыщенных жирных кислот):
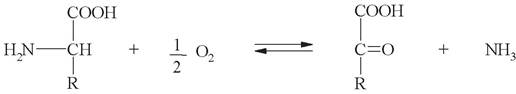
г) окислительное дезаминирование (с образованием кетокислот). У человека окислительное дезаминирование является основным путем катаболизма аминокислот:
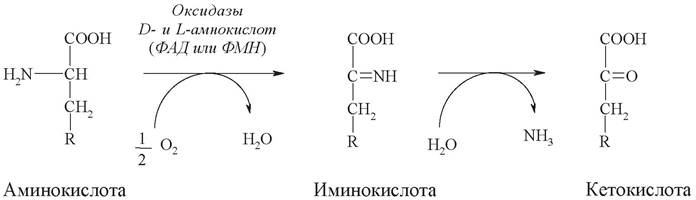
Выделяют два типа окислительного дезаминирования: прямое и непрямое. Если в ходе дезаминирования аминогруппа сразу превращается в аммиак, то реакция называется прямым окислительным дезаминированием. Прямое окислительное дезаминирование может происходить как в аэробных, так и в анаэробных условиях:
1. Аэробное прямое окислительное дезаминирование катализируется оксидазами D-аминокислот и L-аминокислот с участием коферментов ФАД и ФМН соответственно. Реакции, катализируемые оксидазами, в клетках протекают медленно.
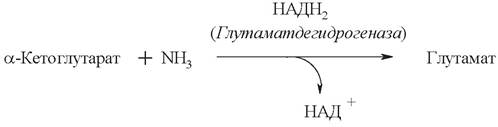
2. Анаэробное прямое окислительное дезаминирование существует только для L-глутаминовой кислоты и катализируется ферментом глутаматдегидрогеназой. Фермент присутствует в митохондриях всех клеток организма (за исключением мышечных). Глутаматдегидрогеназная реакция нуждается в присутствии коферментов НАД+ или НАДФ+:
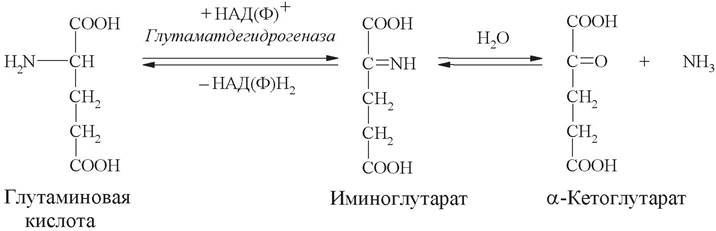
В ходе дезаминирования глутамата аминогруппа сразу превращается в аммиак.
Реакция дезаминирования глутамата ферментом глутаматдегидрогеназой обратима. Обратная реакция - восстановительное аминирование α-кетоглутарата - служит источником глутамата в клетках и обезвреживает аммиак.
Все аминокислоты, кроме глутаминовой, дезаминируются непрямым путем. Непрямое окислительное дезаминирование активно протекает во всех клетках организма и состоит из двух этапов:
1. Трансаминирования с α-кетоглутаратом с образованием глутамата;
2. Прямого окислительного дезаминирования глутамата.
В результате трансаминирования α-кетоглутарат превращается в глутамат, а аминокислоты теряют аминогруппы и превращаются в α-кетокислоты. Далее их углеродный скелет катаболизируется специфическими путями и вовлекается в цикл Кребса, где сгорает до углекислого газа и воды.
Глутаминовая кислота является коллектором всех аминогрупп, и только она подвергается прямому окислительному дезаминированию с образованием аммиака и α-кетоглутаровой кислоты.
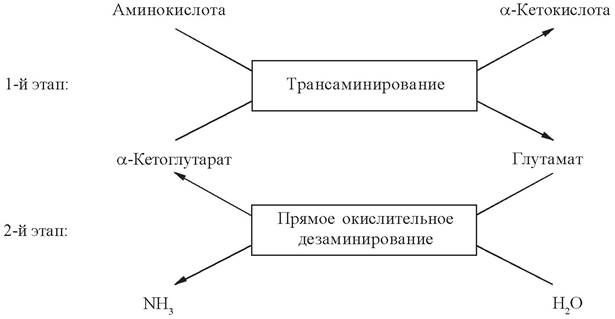
Таким образом, реакция прямого окислительного дезаминирования тесно связана с трансаминированием аминокислот. Сопряжение реакций трансаминирования и дезаминирования направляет поток лишнего аминного азота в печень для синтеза мочевины и в почки для синтеза аммонийных солей.
Декарбоксилирование аминокислот катализируется декарбоксилазами, коферментом которых является пиридоксальфосфат. Продукты декарбоксилирования обладают высокой биологической активностью, и с этим связано их название «Биогенные амины» (табл. 8):
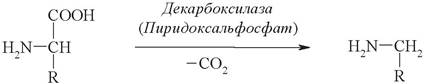
Таблица 8. Биогенные амины
Аминокислота |
Амин |
Функция |
Серин |
Этаноламин |
Составная часть фосфолипидов |
Гистидин |
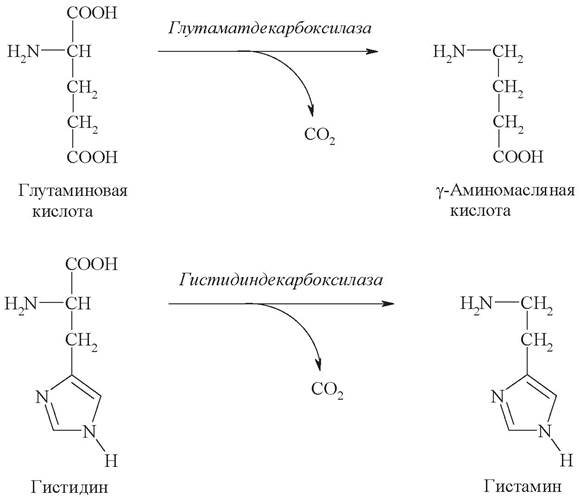
Гистамин |
Образуется в тучных клетках. Является сильным сосудорасширяющим фактором |
Треонин |
Аминопропанол |
Составная часть витамина В12 |
Триптофан |
Серотонин |
Образуется в нейронах гипоталамуса, является возбуждающим нейромедиатором |
Цистеин |
Цистеамин |
Составная часть КоА |
Глутаминовая кислота |
y-Аминомасляная кислота (ГАМК) |
Образуется в ткани головного мозга, является тормозным нейромедиатором |
Тирозин |
Дофамин |
Образуется в почках, надпочечниках, синаптических ганглиях, нервах. Является нейромедиатором двигательных структур головного мозга |
В качестве примера показаны реакции декарбоксилирования с образованием ГАМК и гистамина:
5.4. Механизм токсического действия аммиака
Одним из конечных продуктов обмена аминокислот является высокотоксичный для организма аммиак. Даже небольшое повышение его концентрации оказывает неблагоприятное действие на организм и прежде всего на центральную нервную систему:
1. Аммиак легко проникает через мембраны в клетки и в митохондриях сдвигает реакцию, катализируемую глутаматдегидрогеназой, в сторону образования глутамата:
Уменьшение концентрации α-кетоглутарата вызывает угнетение реакции транс- аминирования аминокислот, а также снижение скорости цикла Кребса и развитие энергодефицита.
2. Аммиак является основанием, и повышение его концентрации в крови сдвигает рН в щелочную сторону, вызывая алкалоз, который увеличивает сродство гемоглобина к кислороду, что препятствует отдаче им кислорода. В результате развиваются гипоксия тканей, энергодефицит, от которого, главным образом, страдает головной мозг.
3. Аммиак является липофильным веществом, он легко проникает в клетки центральной нервной системы. Высокие концентрации аммиака, при участии глутаминсинтетазы, стимулируют синтез глутамина из глутамата в нервной ткани:
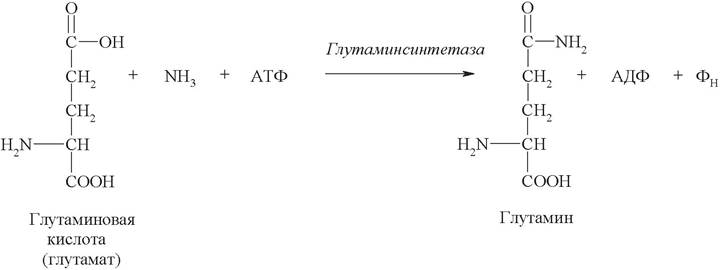
Накопление глутамина в клетках нейроглии приводит к повышению в них осмотического давления, набуханию астроцитов и в больших концентрациях вызывает отек мозга. Снижение концентрации глутамата нарушает обмен аминокислот и нейромедиаторов, в частности, синтез y-аминомасляной кислоты (ГАМК), являющейся основным тормозным медиатором. При недостатке ГАМК и других медиаторов нарушается проведение нервного импульса, возникают судороги.
4. Растворяясь в воде, аммиак образует ион аммония (NH+4), который практически не проникает через цитоплазматические и митохондриальные мембраны. Избыток NH+4 в крови нарушает трансмембранный перенос одновалентных катионов Na+ и К+, конкурируя с ними за ионные каналы, что также влияет на проведение нервных импульсов.
5. Низкие концентрации аммиака стимулируют дыхательный центр, а высокие угнетают.
5.5. Обезвреживание аммиака
5.5.1. Синтез мочевины
Практически весь аммиак удаляется из организма через почки в виде мочевины, которая синтезируется в печени, и в виде солей иона аммония, образующихся в эпителии канальцев почек.
Мочевина - основной конечный продукт азотистого обмена, в составе которого из организма выделяется до 90 % всего выводимого азота. Экскреция мочевины в норме составляет ~ 25 г/сут. При повышении количества потребляемых с пищей белков экскреция мочевины увеличивается. Мочевина синтезируется только в печени.
Цикл мочевинообразования (цикл мочевины, орнитиновый цикл, цикл Кребса - Ганзелейта) - циклический процесс. Он происходит в 3 этапа, включающих 5 реакций, каждая из которых катализируется отдельным ферментом:
- синтез аминокислоты цитруллина (две реакции);
- синтез аминокислоты аргинина (две реакции);
- образование мочевины (одна реакция).
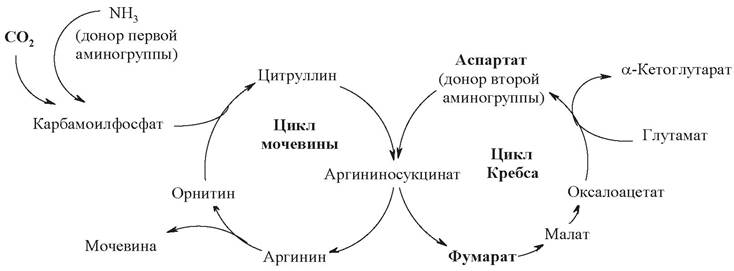
Синтез мочевины начинается в митохондриях (первая и вторая реакции), оставшиеся три реакции идут в цитозоле (рис. 22).
Рис. 22. Цикл мочевинообразования
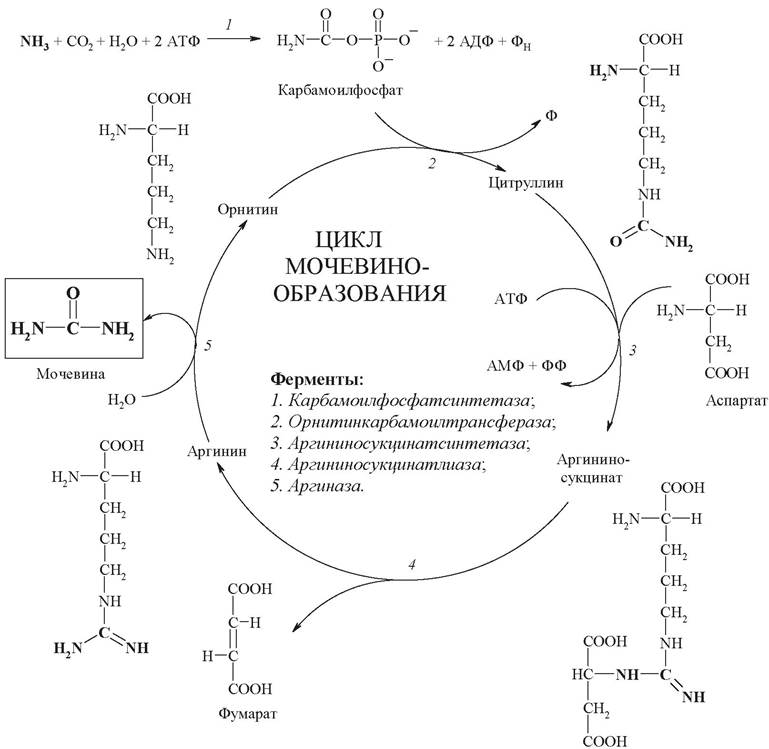
Первый этап - синтез аминокислоты цитруллина протекает в митохондриях печени, где аммиак обезвреживается путем связывания с углекислым газом и образования карбамоилфосфата при участии фермента карбамоилфосфатсинтетазы (1).
Затем следует реакция конденсации образовавшегося карбамоилфосфата и аминокислоты орнитина, катализируемая ферментом орнитинкарбамоилтрансферазой, в ходе реакции образуется цитруллин и регенерирует молекула неорганического фосфата (2).
Второй этап - синтез аргинина из цитруллина и аспартата (донора аминогруппы) протекает уже в цитозоле печени и включает две реакции. Первая - конденсация цитруллина и аспарагиновой кислоты с образованием аргининосукцината катализируется аргининосукцинатсинтетазой (3). Вторая реакция - аргининосукцинат распадается на аргинин и фумаровую кислоту при участии фермента аргининосукцинатлиазы (4).
На третьем этапе аргинин расщепляется на мочевину и орнитин под действием фермента аргиназы (5).
Суммарную реакцию синтеза мочевины можно представить следующим образом:
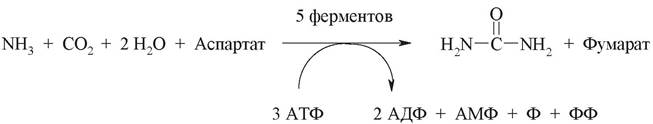
Из анализа цикла мочевины следует, что включение атома азота происходит в двух реакциях. Один из атомов азота поступает в форме NH3 в реакции 1 и является продуктом дезаминирования аминокислот, а другой включается в составе аспартата (реакция 3). Этот второй атом азота может поступать в аспартат из любой аминокислоты путем трансаминирования с оксалоацетатом. Следовательно, атомы азота в мочевине имеют разное происхождение:
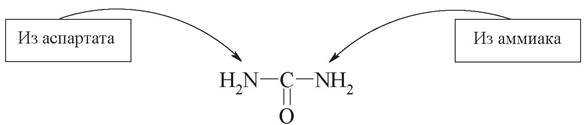
Связь цикла мочевинообразования и цикла Кребса
Между циклом Кребса и циклом мочевинообразования имеются сложные связи, определяющие в известной степени скорость реакций, зависимую от энергетических потребностей клетки и концентраций конечных продуктов метаболизма. Пусковые реакции как цикла мочевины, так и реакции цикла Кребса протекают в митохондриальном матриксе.
Синтез мочевины связан с циклом Кребса тремя основными путями:
1. Фумаpат является общим метаболитом цикла мочевины и цикла Кребса. В цикле Кребса фумаpат превращается в малат и затем в оксалоацетат. Оксалоацетат подвергается трансаминированию с образованием аспартата, который вступает в цикл синтеза мочевины. Оксалоацетат является важным метаболитом, участвующим в ЦТК и глюконеогенезе.
2. При окислении малата до оксалоацетата в цикле Кребса образуется 3 молекулы АТФ, которые в дальнейшем используются для синтеза мочевины.
3. Поступление СО2, необходимого для образования мочевины, обеспечивается работой цикла Кребса:
Таким образом, при помощи этого необычного сцепленного механизма происходит переплетение реакций обоих циклов. Этот механизм получил название «велосипед Кребса» или «двухколесный велосипед Кребса».
5.5.2. Синтез аммонийных солей
Непосредственный синтез аммонийных солей происходит в просвете канальцев почек из секретируемых сюда аммиака и ионов водорода и фильтрующихся органических (уксусной, щавелевой, молочной кислот) и неорганических (фосфаты, хлориды, сульфаты) анионов первичной мочи. Около 10 % всего аммиака выводится почками в виде аммонийных солей.
5.6. Судьба безазотистого остатка аминокислот
Катаболизм углеродных скелетов, полученных в результате дезаминирования аминокислот, приводит к образованию либо ацетил-КоА, а далее из него жиров или кетоновых тел (кетогенные аминокислоты), или к образованию метаболитов, способных включаться в глюконеогенез (глюкогенные аминокислоты).
Существует несколько специфических путей катаболизма аминокислот, которые сходятся к 6 продуктам, которые вступают в цикл Кребса, где полностью окисляются до СО2 и Н2О.
К глюкогенным относятся аминокислоты (аланин, глицин, треонин, серин, цистеин, метионин, аспартат, аспарагин, глутамат, глутамин, аргинин, пролин, гистидин), при распаде которых образуются субстраты глюконеогенеза - пируват или метаболиты цикла Кребса (оксалоацетат, α-кетоглутарат, сукцинал-КоА).
Строго кетогенными аминокислотами являются лизин и лейцин, при их окислении образуется исключительно ацетил-КоА, который далее принимает участие в синтезе кетоновых тел, жирных кислот и холестерола.
Выделяют небольшую группу смешанных аминокислот, из которых образуются как пируват или метаболиты цикла Кребса, так и ацетил-КоА. Смешанными аминокислотами являются фенилаланин, тирозин, изолейцин, триптофан.
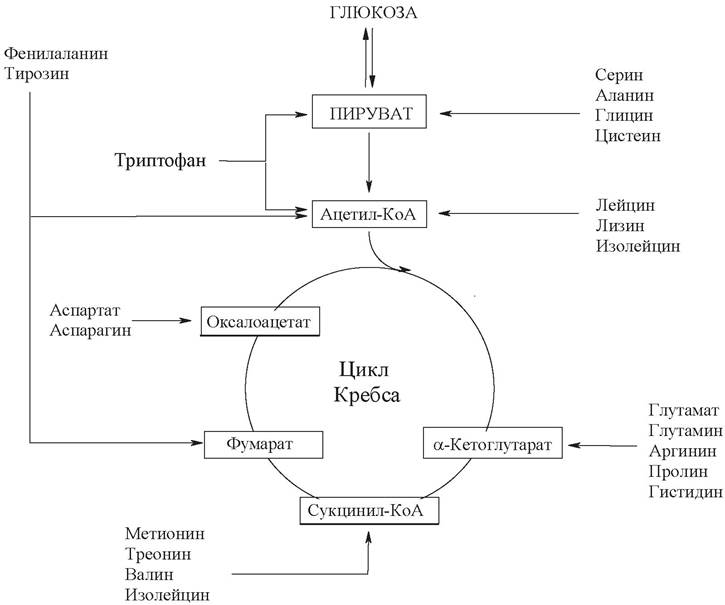
Для ряда аминокислот существуют специфические пути обмена, обусловленные особенностями их строения и физиологической роли в организме.
5.7. Биосинтез аминокислот
Растения и многие виды бактерий содержат ферменты, необходимые для синтеза всех необходимых α-кетокислот. Животные утратили способность синтезировать некоторые α-кетокислоты, из которых могут быть получены незаменимые аминокислоты. Другие α-кетокислоты могут образовываться в результате метаболизма иных соединений (в основном из глюкозы) и использоваться для синтеза заменимых аминокислот.
Человек и животные способны синтезировать из неаминокислотных предшественников только 8 из 20 аминокислот, необходимых для синтеза белка (заменимые аминокислоты), 2 аминокислоты (частично заменимые аминокислоты) синтезируются в ограниченных количествах и еще 2 образуются из незаменимых аминокислот (условно заменимые аминокислоты). Углеродный скелет аминокислот образуется из промежуточных метаболитов гликолиза, пентозофосфатного пути, цикла Кребса. Основными путями биосинтеза аминокислот являются: прямое аминирование α-кетокислот или ненасыщенных органических кислот, реакции трансаминирования, ферментативные взаимопревращения отдельных аминокислот - как заменимых, так и незаменимых.
Ниже приведена схема синтеза одиннадцати заменимых аминокислот, углеродный скелет которых может образовываться из метаболитов гликолиза и цикла Кребса:
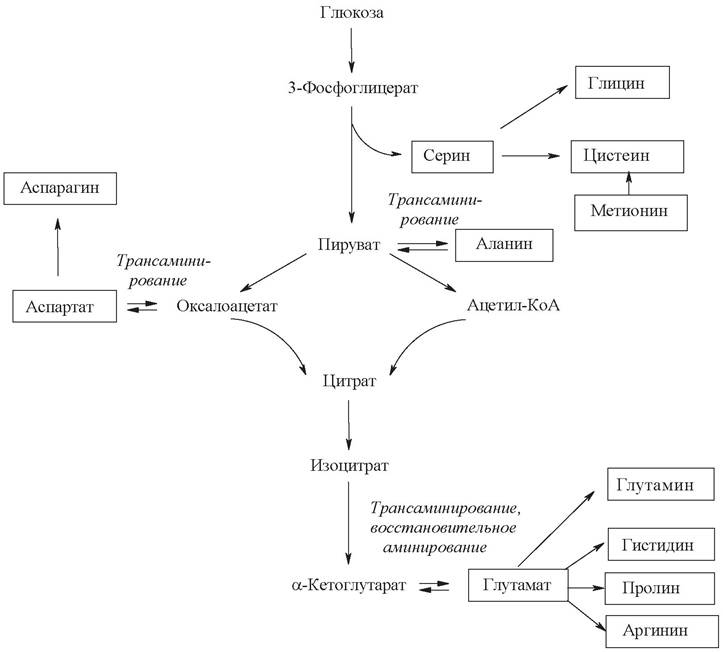
Необходимо отметить, что источником атома серы в молекуле условно заменимого цистеина служит незаменимый метионин. Двенадцатая аминокислота - тирозин - синтезируется путем гидроксилирования незаменимой аминокислоты фенилаланина и на схеме не показана.
5.8. Переваривание и всасывание нуклеиновых кислот
С пищей в сутки в организм человека поступает около 1 г нуклеиновых кислот. Переваривание нуклеиновых кислот происходит в тонкой кишке (рис. 23). Нуклеиновые кислоты пищи подвергаются гидролизу под действием ферментов панкреатического сока. Ферменты, катализирующие распад нуклеиновых кислот, называют нуклеазами или фосфодиэстеразами. По специфичности действия различают дезоксирибонуклеазы (ДНКазы, расщепляющие ДНК) и рибонуклеазы (РНКазы, расщепляющие РНК).
Рис. 23. Схема расщепления (гидролиза) нуклеиновых кислот
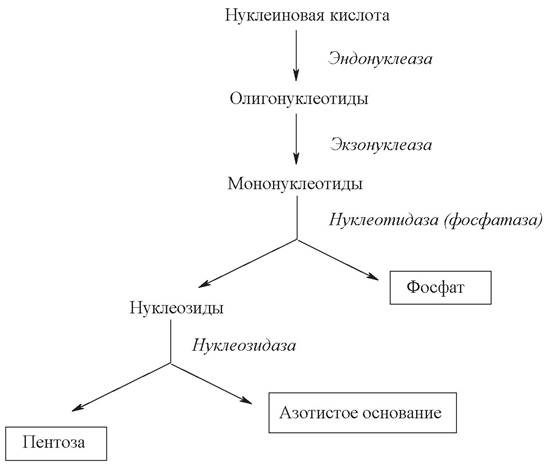
Различают эндонуклеазы и экзонуклеазы. Эндонуклеазы действуют на внутренние фосфодиэфирные связи в молекулах ДНК и РНК и обеспечивают распад нуклеиновых кислот в основном до олигонуклеотидов. Экзонуклеазы отщепляют нуклеотиды с 3- или 5-конца полинуклеотидной цепи и приводят к образованию свободных мононуклеотидов.
В результате действия внутриклеточных эндо- и экзонуклеаз нуклеиновые кислоты расщепляются до мононуклеотидов, которые при участии ферментов тонкого кишечника нуклеотидаз (фосфатаз) гидролизуются с образованием соответствующего нуклеозида и ортофосфорной кислоты.
Нуклеозиды далее расщепляются под действием ферментов нуклеозидаз до азотистых оснований и пентоз (рибозы или дезоксирибозы).
Продукты переваривания нуклеиновых кислот поступают в кровь, а затем в печень и другие органы. В клетках организмов обмен РНК протекает значительно более интенсивно, чем обмен ДНК.
В конечном итоге нуклеиновые кислоты расщепляются на азотистые основания, пентозы и фосфорную кислоту.
Азотистые основания превращаются в конечные продукты обмена и выводятся с мочой. Пуриновые азотистые основания в процессе катаболизма теряют аминогруппу в виде аммиака, окисляются и превращаются в мочевую кислоту. Пиримидиновые основания подвергаются более глубокому расщеплению до углекислого газа, воды и аммиака.
Пентозы вовлекаются в пентозофосфатный путь и могут быть окислены до углекислого газа и воды.
Фосфорная кислота распаду не подвергается и используется для фосфорилирования органических соединений или выводится из организма с мочой.
Продукты гидролиза нуклеиновых кислот поступают в клетки организма и используется для синтеза нуклеотидов, нуклеиновых кислот, а также удовлетворения энергетических потребностей организма.
5.9. Катаболизм пуриновых оснований
Реакции расщепления пуриновых нуклеотидов - аденозинмонофосфата (АМФ) и гуанозинмонофосфата (ГТФ) - происходят различными путями, но приводят к одному продукту - ксантину (рис. 24). Первоначально нуклеотиды подвергаются реакции гидролитического отщепления фосфатного остатка с образованием соответствующих нуклеозидов: аденозина и гуанозина. Затем гуанозин гидролизуется до свободного основания гуанина, которое непосредственно превращается в ксантин. Образование ксантина из аденозина происходит через ряд промежуточных стадий: дезаминирование аденозина с образованием инозина, гидролиз инозина до свободного основания гипоксантина и, наконец, окисление гипоксантина до ксантина. Превращение гипоксантина в ксантин происходит под действием фермента ксантиноксидазы. Этот фермент в значительных количествах обнаруживается в печени и кишечнике и окисляет пурины молекулярным кислородом, а в качестве побочного продукта образует высокотоксичный супероксид-радикал (О2-).
Рис. 24. Реакции расщепления пуриновых нуклеотидов
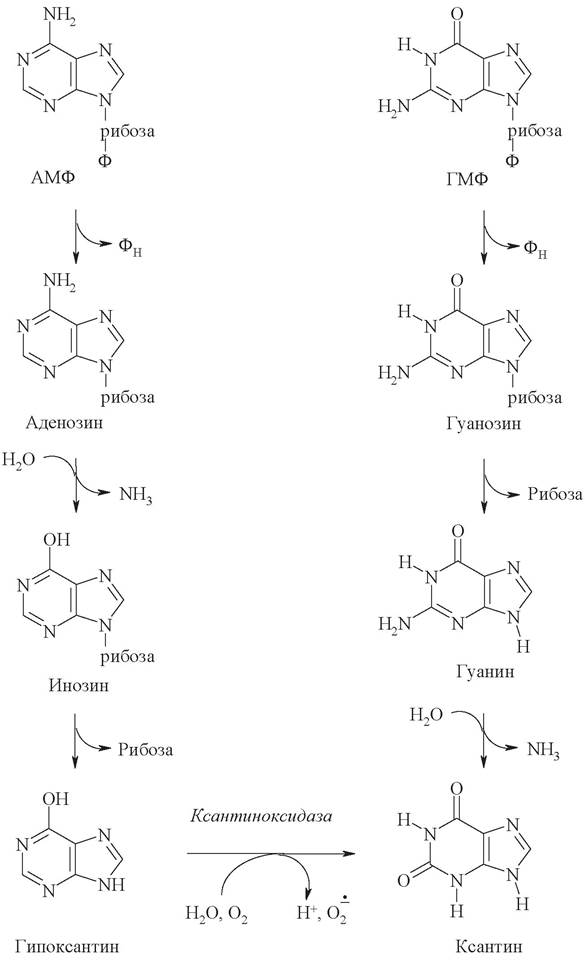
Дальнейшая судьба ксантина может быть различной и зависит от конкретного организма. У человека, как и у большинства приматов, птиц, некоторых рептилий и большинства насекомых, ксантин под действием ксантиноксидазы превращается в мочевую кислоту. Образование мочевой кислоты происходит преимущественно в печени. В сутки в организме человека образуется 0,5-1 г мочевой кислоты, которая удаляется из организма главным образом с мочой и незначительно с фекалиями. Мочевая кислота плохо растворяется в воде, и при нарушении обмена нуклеиновых кислот мочевая кислота откладывается в виде кристалликов в суставах пальцев, хрящах, в коже и мышцах, образуя узелки. Вокруг узелков развивается болезненный очаг воспаления. С этими симптомами связано заболевание подагра.
У всех остальных наземных животных конечным продуктом катаболизма пуринов является более растворимый аллантоин, который образуется в ходе дальнейшего окисления мочевой кислоты.
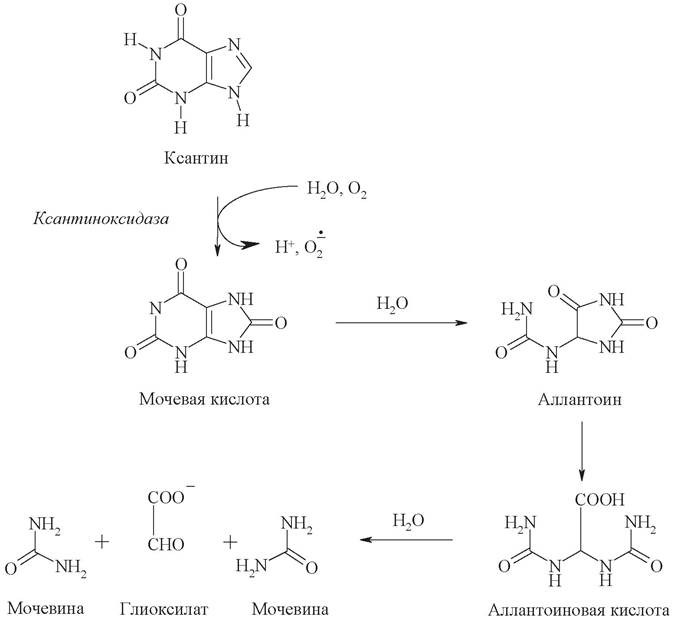
У некоторых животных аллантоин может распадаться далее до мочевины и аммиака. У амфибий и рыб аллантоин гидролизуется до аллантоиновой кислоты. Во многих организмах аллантоиновая кислота расщепляется до мочевины и глиоксилата.
5.10. Катаболизм пиримидиновых оснований
Основной путь катаболизма пиримидиновых оснований, образующихся в результате гидролиза пиримидиновых нуклеотидов, протекает через восстановление урацила и тимина до полностью гидрированных структур - дигидроурацила и дигидротимина соответственно. Цитозин также расщепляется по этому пути после его дезаминирования до урацила (рис. 25).
Рис. 25. Катаболизм пиримидиновых оснований
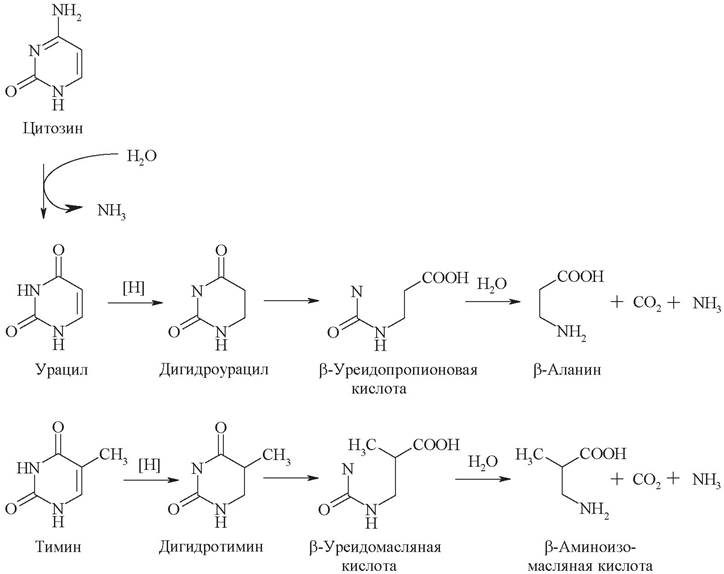
Раскрытие кольца в дигидроурациле и последующий гидролиз приводят к образованию β-аланина, углекислого газа и аммиака. Аналогичные реакции при расщеплении тимина дают β-аминоизомасляную кислоту, углекислый газ и аммиак.
Продукты катаболизма пиримидинов выводятся из организма или повторно используются в других метаболических процессах. Так, аммиак включается в орнитиновый цикл образования мочевины. β-аланин используется микрофлорой кишечника при биосинтезе пантотеновой кислоты (витамин В3), которая, в свою очередь, необходима для синтеза коэнзима А и ацилпереносящего белка, участвующего в синтезе жирных кислот. β-аминоизомасляная кислота участвует в реакциях трансаминирования с α-кетоглутаратом с образованием метималонового полуальдегида, который используется в синтезе пропионата и сукцината.
5.11. Анаболизм нуклеотидов
Почти все организмы способны синтезировать пиримидиновые и пуриновые нуклеотиды из простых соединений, например, из СО2, NH3, аспартата, глицина, глутамина и рибозы.
В синтезе обоих типов нуклеотидов фосфорибозильный фрагмент переносится в виде 5-фосфорибозил-1-пирофосфата (ФРПФ), который образуется при фосфорилировании рибозо-5-фосфата - промежуточного метаболита пентозофосфатного пути:
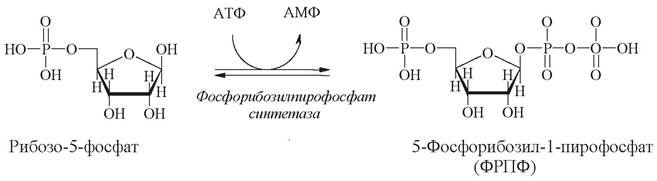
Для синтеза пуриновых и пиримидиновых нуклеотидов существуют отдельные пути. Метаболические пути, ведущие к образованию пуриновых и пиримидиновых нуклеотидов, различаются в основном тем, на каком этапе синтеза возникает β-гликозидная связь (табл. 9). При синтезе пуринов эта связь образуется на первом этапе, и циклическая система строится уже после того, как связь образовалась. В отличие от этого синтез пиримидинового кольца завершается еще до образования связи между этим кольцом и рибозо-5-фосфатом.
Таблица 9. Сравнительная характеристика путей синтеза пуриновых и пиримидиновых нуклеотидов
Особенности синтеза |
Путь синтеза |
|
Пуринов |
Пиримидинов |
|
Последовательность синтеза |
1. Образование N-гликозидной связи. 2. Сборка кольцевой структуры |
1. Сборка кольцевой структуры. 2. Образование N-гликозидной связи |
Ключевая реакция |
Образование фосфорибозиламина (фосфорибозиламидотрансфераза) |
Образование карбамоилфосфата (карбамоилфосфатсинтетаза) |
Локализация в клетке |
Цитозоль |
Митохондрии и цитозоль |
Регуляция |
Торможение ИМФ, АМФ и ГМФ на нескольких уровнях |
Торможение УТФ карбамоилфосфатсинтетазы |
Первым продуктом нуклеотидной природы пуринового пути является инозин- 5-монофосфат (ИМФ), пиримидинового пути - уридинмонофосфат (УМФ). Из ИМФ и УМФ получаются все остальные пуриновые и пиримидиновые нуклеотиды соответственно.
5.11.1. Биосинтез пуриновых мононуклеотидов
Биосинтез первого пуринового нуклеотида - ИМФ - включает 10 ферментативных реакций и протекает с затратой энергии АТФ (рис. 26). Формирование пуринового кольца начинается сразу на рибозо-5-фосфате, к которому присоединяются атомы азота и углерода. Источниками этих атомов являются аминокислоты глицин, глутамин, аспарагиновая кислота. Часть атомов углерода поставляется коферментами, содержащими в своем составе фолиевую кислоту и биотин. В результате последующих реакций происходит последовательная сборка пуринового кольца на этой основе.
Рис. 26. Биосинтез пуриновых мононуклеотидов
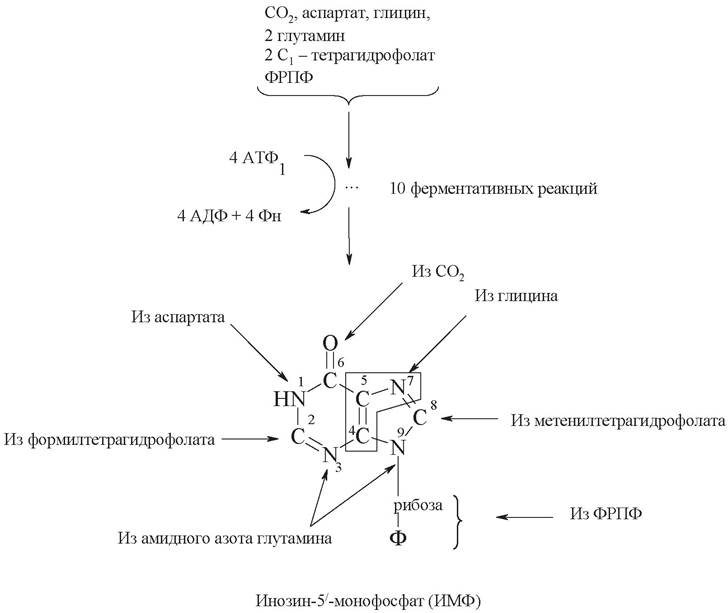
Происхождение каждого атома пуринового гетероцикла установлено экспериментально с использованием изотопов. Как видно, два атома азота (N3 и N9) пуринового кольца происходят из амидной группы глутамина, третий атом азота (N1) - из аспартата, и, наконец, четвертый (N7) - из глицина. Из глицина происходят также четвертый и пятый атомы углерода. Атомы углерода (С2 и С8) происходят из формиата, а шестой атом углерода - из СО2.
Из ИМФ образуются другие нуклезидмонофосфаты, например, АМФ и ГМФ.
Существует альтернативный путь синтеза пуриновых нуклеотидов. Это обусловлено тем, что на синтез циклической структуры пуринов затрачивается значительное количество энергии в форме АТФ. Поэтому в клетке содержатся ферменты, которые используют («реутилизируют») пуриновые основания, образующиеся при катаболизме нуклеотидов, до их превращения в ксантин и мочевую кислоту:
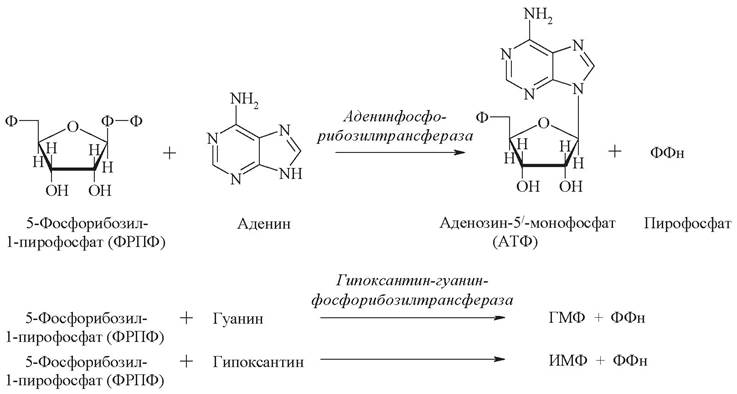
Ферменты аденинфосфорибозилтрансфераза и гипоксантингуанинфосфорибозилтрансфераза утилизируют свободные пурины, превращая их снова в нуклеотиды при взаимодействии с ФРПФ.
5.11.2. Биосинтез пиримидиновых мононуклеотидов
Биосинтез пиримидиновых мононуклеотидов происходит поэтапно и включает 6 ферментативных реакций (рис. 27). В синтезе уридинмонофосфата циклическая пиримидиновая структура формируется до присоединения к рибозо-5-фосфату.
Рис. 27. Биосинтез пиримидиновых мононуклеотидов
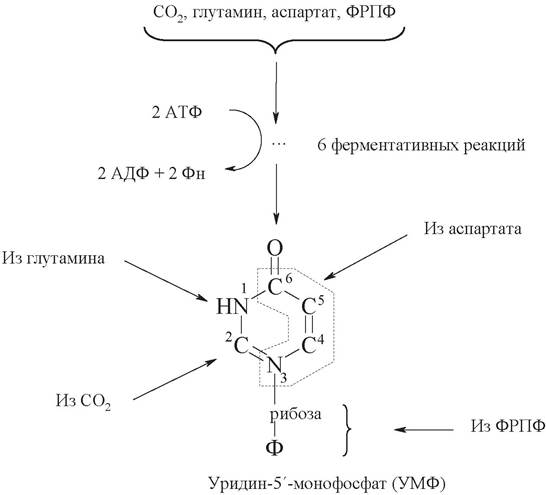
На первом этапе из СО2, глутамина и АТФ образуется карбамоилфосфат. Затем из карбомоилфосфата и аспарагиновой кислоты синтезируется необычное азотистое основание - оротовая кислота, которая содержит пиримидиновое кольцо. Оротовая кислота присоединяется к рибозо-5-фосфату и возникает пиримидиновый нуклеотид оротидинмонофосфат. Далее оротовая кислота в составе нуклеотида преобразуется в уридин пиримидиновые нуклеотиды.
5.11.3. Биосинтез нуклеозидтрифосфатов
Нуклеозидтрифосфаты образуются при участии АТФ из нуклеотид-монофосфатов (НМФ) в результате двух последовательно протекающих реакций фосфорилирования:
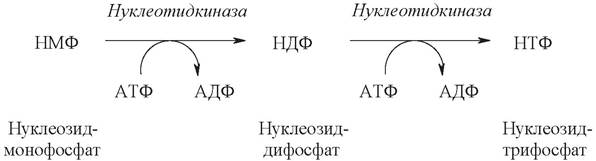
Уридинмонофосфат (УМФ) служит предшественником для трифосфонуклеотидов УТФ и ЦТФ. Причем, образование УТФ происходит в описанные выше две стадии фосфорилирования, а ЦТФ образуется из УТФ путем аминирования в положении 4 пиримидинового кольца.
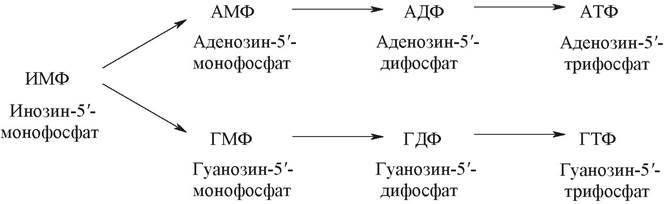
Предшественником ГТФ и АТФ является инозин-5'-монофосфат (ИМФ). В обоих случаях сначала модифицируется гипоксантиновое кольцо ИМФ с образованием соответствующих монофосфонуклеотидов ГМФ и АМФ, которые затем фосфорилируются.
5.11.4. Биосинтез дезоксирибонуклеотидов
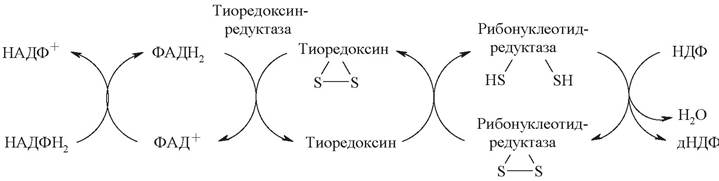
Дезоксирибонуклеотиды образуются из соответствующих рибонуклеозидфосфатов путем восстановления входящей в них рибозы в дезоксирибозу при участии фермента рибонуклеотид-редуктазы.
Источником восстановительных эквивалентов является термостабильный белок - тиоредоксин, который содержит две свободные SH-группы. Тиоредоксинредуктаза катализирует НАДФН-зависимое восстановление тиоредоксина (рис. 28).
Рис. 28. Биосинтез дезоксирибонуклеотидов
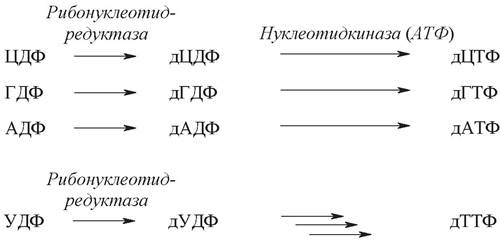
Если в реакцию вступают нуклеозиддифосфаты, то синтезируются дезоксирибонуклеотиды: дУДФ, дЦДФ, дГДФ и дАДФ. Дезоксирибонуклеозиддифосфаты дЦДФ, дГДФ и дАДФ далее могут фосфорилироваться до соответствующих дезоксирибонуклеозидтрифосфатов дЦТФ, дГТФ и дАТФ. дУДФ служит предшественником дТТФ.
Биосинтез всех дезоксирибонуклеозид-5'-трифосфатов и рибонуклеозид-5'- трифосфатов регулируется в клетке таким образом, что они образуются зависимо друг от друга в строго определенных соотношениях.
5.12. Синтез нуклеиновых кислот
Для синтеза нуклеиновых кислот используются мононуклеотиды обязательно в трифосфатной форме. Такие нуклеотиды содержат в своей молекуле три остатка фосфорной кислоты и обладают повышенным запасом энергии. Для синтеза РНК используются АТФ, ГТФ, УТФ, ЦТФ, а для синтеза ДНК, соответственно, дАТФ, дГТФ, дТТФ, дЦТФ.
Процесс репликации, или редупликации, ДНК иначе можно назвать удвоением. Он происходит перед делением клетки. Для реализации репликации необходимы: матрица - расплетенная цепь ДНК; субстраты, участвующие в полимеризации ДНК (дезоксирибонуклеозидтрифосфаты); ферменты, катализирующие этот процесс; ионы Mg2+, а также белковые факторы, обеспечивающие деспирализацию двухнитевой ДНК. Синтез ДНК осуществляет фермент ДНК-полимераза. Этот фермент достраивает к каждой из них комплементарную ей цепь. Таким образом, из одной молекулы образуются две одинаковые дочерние молекулы, причем обе цепи ДНК служат матрицами для дочерних цепей. Процесс является полуконсервативным, так как по завершении репликации каждая дочерняя молекула ДНК содержит одну родительскую и одну вновь синтезированную цепь. По мере присоединения к матрице нуклеотиды связываются в полинуклеотидные нити, которые сразу же закручиваются в двойную спираль. Биологический смысл репликации состоит в том, что из одной молекулы ДНК появляются две ее полные копии. Процесс этот идет с очень высокой точностью - ошибки крайне редки.
Процесс синтеза РНК на ДНК-матрице называется транскрипцией. Транскрипцию осуществляет фермент РНК-полимераза. Этот фермент соединяет между собой рибонуклеотиды, составляющие остов молекулы РНК. Делает это фермент на основании считывания последовательности молекулы ДНК и достраивания комплементарной ей последовательности. Показано, что в этом процессе только одна из двух цепей ДНК играет роль матрицы.
Вопросы для проверки усвоения материала
1. Аминокислоты: определение, биологическое значение. Классификация аминокислот по строению, полярности боковых радикалов, физиологическим свойствам. Изомерия аминокислот, биологическое значение. Растворимость аминокислот и кислотно-основные свойства, понятие об изоэлектрической точке.
2. Биологические функции белков: ферментативная, структурная, регуляторная, защитная, рецепторная, транспортная, двигательная, энергетическая. Особенности строения структурных (коллаген, кератин, фиброин) и транспортных (гемоглобин) белков.
3. Нуклеиновые кислоты. Нуклеотиды и нуклеозиды, строение, биологические функции. Важнейшие представители мононуклеотидов и олигонуклеотидов. ДНК и РНК: строение, биологические функции.
4. Пути обмена аминокислот в клетке: реакции декарбоксилирования, переаминирования и дезаминирования, биологическое значение. Судьба безазотистого остатка аминокислот, глюкогенные и кетогенные аминокислоты. Использование аминокислот для синтеза биологически активных веществ.
5. Образование, токсичность и обезвреживание аммиака. Цикл мочевины: реакции, регуляция, биологическое значение. Взаимосвязь обмена аминокислот с циклом Кребса, обменом углеводов и липидов.
6. Этапы биосинтеза белка (рекогниция, транскрипция, трансляция): сущность процессов, субстраты, ферменты, локализация в клетке. Катаболизм белка в лизосомах и протеасомах, механизмы и биологическое значение. Роль гормонов и нервной системы в регуляции белкового обмена.
Письменное домашнее задание
Обязательное
1. Переваривание белков в желудке обеспечивает фермент желудочного сока пепсин. Главный протеолитический фермент сока поджелудочной железы - трипсин - продолжает переваривание в двенадцатиперстной кишке. Известно, что пепсин преимущественно гидролизует пептидные связи, образованные аминогруппами ароматических аминокислот, а трипсин - карбоксильными группами основных аминокислот. На какие фрагменты будет расщеплен полипептид следующего строения
H2N-Ala-Gly-Phe-Leu-Ser-Lys-Ala-Val-Ile-Met-Arg-Ser-Glu-Tyr-Gly-COOH при действии пепсина, трипсина и последовательном действии обоих ферментов?
2. Напишите химические реакции обмена аминокислот в клетках и дайте названия катализирующим их ферментам:
а) декарбоксилирование гистидина;
б) окислительное дезаминирование глутамата;
в) переаминирование глутамата и оксалоацетата;
г) гидроксилирование фенилаланина с образованием тирозина;
д) декарбоксилирование триптофана;
е) гидролиз аргинина с образованием мочевины.
Какие витамины являются предшественниками коферментов, участвующих в этих реакциях? Какие из приведенных аминокислот являются незаменимыми?
3. В эксперименте на животных изучали метаболизм аминокислот. Для этого им скармливали аспарагиновую кислоту, меченую радиоактивным азотом по аминогруппе. Через некоторое время в моче животных радиоактивная метка была обнаружена в двух низкомолекулярных веществах. Первое вещество оказалось хорошо растворимо в воде, второе легко образовывало осадок. Эксперимент повторили в условиях белкового голодания. Выведение радиоактивных продуктов обмена с мочой резко увеличилось. О каких веществах идет речь в задаче? Как объяснить наблюдавшиеся закономерности? В каких еще веществах мог быть обнаружен меченый атом азота?
4. Напишите структурную формулу участка молекулы ДНК, содержащую азотистые основания А - Г - Ц, и комплементарного ему участка. Какие типы связи образуются между нуклеотидами в пределах одной цепи и между двумя комплементарными цепями?
Дополнительное
1. Напишите химические реакции обмена аминокислот в клетках и дайте названия катализирующим их ферментам:
а) декарбоксилирование цистеина;
б) внутримолекулярное дезаминирование гистидина;
в) переаминирование лейцина и α-кетоглутарата;
г) гидроксилирование триптамина с образованием серотонина;
д) метилирование норадреналина в адреналин;
е) окисление аргинина в цитруллин с образованием оксида азота (NO).
2. В медицине используются лекарственные препараты, влияющие на обмен нуклеотидов: оротат калия, инозин (рибоксин), 5-фторурацил. Напишите их структурные формулы. Укажите, какие метаболические пути они активируют или ингибируют, каким лечебным действием обладают.
Типовой вариант тестового контроля по теме «Обмен аминокислот, белков и нуклеиновых кислот»
Инструкция: при отсутствии дополнительных указаний в вопросе теста выберите один верный вариант ответа.
1. Выберите название фермента, катализирующего следующую реакцию:
а) карбамоилфосфатсинтетаза;
б) аргининосукцинатсинтетаза;
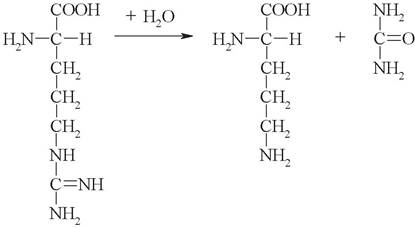
в) аргининосукцинатлиаза;
г) аргиназа;
д) уреаза.
2. Укажите продукты декарбоксилирования аминокислот:
а) новая аминокислота и кетокислота;
б) углекислый газ и вода;
в) новая аминокислота и аммиак;
г) углекислый газ и амин;
д) аммиак и вода.
3. Выберите фермент, катализирующий окислительное дезаминирование аминокислот:
а) аспартатаминотрансфераза;
б) аланинаминотрансфераза;
в) глутаминаза;
г) глутаматдегидрогеназа;
д) глутаматдекарбоксилаза.
4. Углеродные скелеты аминокислоты АСП и АСН превращаются в субстрат цикла Кребса оксалоацетат, поэтому данные аминокислоты называют:
а) условно заменимыми;
б) незаменимыми;
в) глюкогенными;
г) кетогенными;
д) протеиногенными.
5. Укажите свойства мочевины, делающие ее нетоксичной:
а) сильное основание, растворима в воде, легко выводится с мочой;
б) слабое основание, растворима в воде, легко выводится с мочой;
в) сильное основание, растворима в липидах, легко выводится с мочой;
г) слабое основание, растворима в липидах, не выводится с мочой;
д) слабое основание, растворима в воде, не выводится с мочой.
6. Участок молекулы ДНК, комплементарный участку ААГЦЦТ, имеет вид:
а) ААГЦЦТ;
б) ТТАГГУ;
в) ТТЦГАА;
г) ЦЦАГУУ;
д) ТАГЦТУ.
7. Какая связь образуется между двумя фосфорными остатками в нуклеотиде:
а) пептидная;
б) фосфоангидридная;
в) фосфодиэфирная;
г) гликозидная;
д) водородная.
8. Какие процессы в биосинтезе белка происходят на этапе транскрипции:
а) сборка рибосом и синтез полипептидной цепи;
б) синтез и созревание мРНК;
в) взаимодействие тРНК с аминокислотой;
г) присоединение небелкового компонента к полипептидной цепи;
д) ничего из перечисленного.
9. Вставьте пропущенные слова (3 ответа) в предложение: «Мочевая кислота - конечный продукт распада ... нуклеотидов, ... растворима в воде и может образовывать ... в почках».
10. Напишите названия и номера классов ферментов (6 ответов), катализирующих реакции 1, 2 и 3: