БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 4. БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ И БЕЛКОВ (МАТРИЧНЫЕ БИОСИНТЕЗЫ). ОСНОВЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИ
II. Репликация
Живые организмы в течение S-фазы клеточного цикла, которая предшествует делению клетки, удваивают содержание ДНК таким образом, что каждая дочерняя клетка после деления получает набор хромосом, идентичный родительской клетке. Процесс удвоения хромосом называют репликацией (редупликацией).
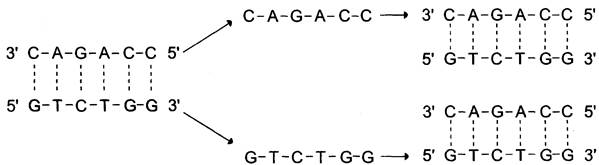
Хромосома содержит одну непрерывную двухцепочечную молекулу ДНК. При репликации каждая цепь родительской двухцепочечной ДНК служит матрицей для синтеза новой комплементарной цепи. Вновь образованная двойная спираль имеет одну исходную (родительскую) и одну вновь синтезированную (дочернюю) цепь. Такой механизм удвоения ДНК получил название «полуконсервативная репликация» (рис. 4-14). Первичная структура дочерней цепи определяется первичной структурой родительской цепи, потому что в основе её образования лежит принцип комплементарности оснований (G = С и А = Т).
Рис. 4-14. Полуконсервативная репликация.
Ферменты и белки, участвующие в репликации, должны работать быстро и точно. Эти условия выполняются с помощью особого мультиферментного комплекса.
Репликацию можно разделить на 4 этапа: образование репликативной вилки (инициация), синтез новых цепей (элонгация), исключение праймеров, завершение синтеза двух дочерних цепей ДНК (терминация).
А. Иницифция репликации
Синтез ДНК у эукариотов происходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы — факторы роста.Факторы роста связываются рецепторами мембран клеток, которые передают сигнал, побуждающий клетку к началу репликации (см. раздел 11).
Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях.
В образовании репликативной вилки принимает участие ряд белков и ферментов. Так, семейство ДНК-топоизомераз (I, II и III), обладая нуклеазной активностью, участвует в регуляции суперспирализации ДНК. Например, ДНК-топоизомераза I разрывает фосфоэфирную связь в одной из цепей двойной спирали и ковалентно присоединяется к 5'-концу в точке разрыва (рис. 4-15). По окончании формирования репликативной вилки фермент ликвидирует разрыв в цепи и отделяется от ДНК.
Рис. 4-15. Участие ДНК-топоизомеразы I в образовании репликативной вилки. 1 — фермент расщепляет одну цепь ДНК; между остатком тирозина молекулы фермента и фосфорным остатком цепи образуется ковалентная связь; 2 — происходит локальное раскручивание двойной спирали при участии ДНК-хеликазы; ДНК-топоизомераза I восстанавливает фосфоэфирную связь.
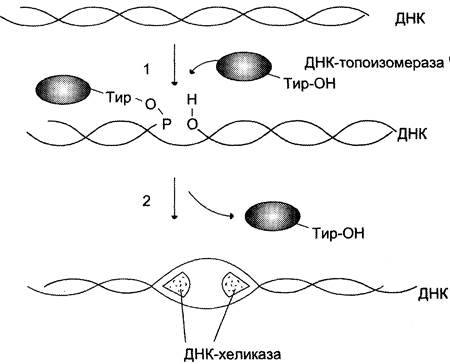
Разрыв водородных связей в двухцепочечной молекуле ДНК осуществляет ДНК-хеликаза. Фермент ДНК-хеликаза использует энергию АТФ для расплетения двойной спирали ДНК.
В результате происходит раскручивание участка суперспирализованной молекулы ДНК. В поддержании этого участка ДНК в раскрученном состоянии участвуют SSB-белки (от англ. single strand binding proteins, т. е. белки, связывающиеся с одноцепочечными нитями ДНК). SSB-белки, не закрывая азотистых оснований, связываются с одноцепочечной ДНК по всей длине разделившихся цепей и таким образом предотвращают их комплементарное скручивание и образование «шпилек». Они обладают большим сродством к одноцепочечным участкам ДНК, независимо от первичной структуры цепей.
Б. Элонгация
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами (рис. 4-16). Субстратами и источниками энергии для синтеза продукта служат 4 макроэргических соединения — дезоксирибонуклеозидтрифосфаты дАТФ, дГТФ, дЦТФ и дТТФ, для активации которых необходимы ионы магния. Нейтрализуя отрицательный заряд нуклеотидов, они повышают их реакционную способность. Ферменты проявляют каталитическую активность только в присутствии предварительно раскрученной матричной двухцепочечной ДНК. Синтез цепей ДНК происходит в направлении 5' —> 3' растущей цепи, т. е. очередной нуклеотид присоединяется к свободному 3'-ОН-концу предшествующего нуклеотидного остатка. Синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, представляющие собой копии матричных цепей.
Рис. 4-16. Репликация.
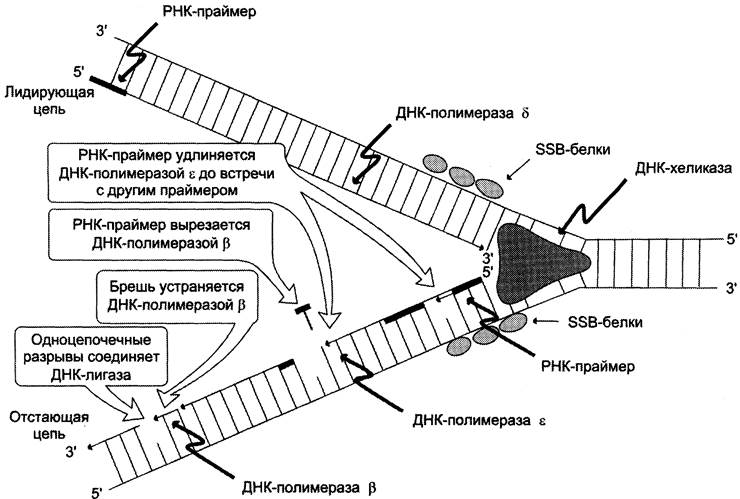
В синтезе эукариотических ДНК принимают участие 5 ДНК-полимераз (α, β, y, δ, ε). ДНК- полимеразы различают по числу субъединиц, молекулярной массе, ассоциации с разными вспомогательными белками, ускоряющими процесс биосинтеза ДНК, и функциональному назначению. ДНК-полимеразы α (альфа), β (бета), δ (дельта), ε (эпсилон) участвуют в синтезе ДНК в ядре клеток, ДНК-полимераза у (гамма) — в репликации митохондриальной ДНК.
ДНК-полимеразы β, δ, ε не могут инициировать образование дочерних цепей, так как не имеют сродства к одиночной нити ДНК. Инициирует репликацию ДНК-полимераза а, которая комплементарна определённому сайту одноцепочечной ДНК. Присоединяясь к нему, ДНК-полимераза а синтезирует небольшой фрагмент РНК — праймер, состоящий из 8 — 10 рибонуклеотидов. ДНК-полимераза а состоит из четырёх субъединиц. Каждая из субъединиц фермента выполняет определённую функцию: «узнавание» сайта репликации, синтез праймера (8 — 10 рибонуклеотидов), синтез фрагмента цепи ДНК, около 50 дезоксирибонуклеотидов. Таким образом, ДНК-полимераза а синтезирует олигонуклеотид, содержащий примерно 60 нуклеотидных остатков; первые 8 — 10 представлены рибонуклеотидами (праймер), а остальные — дезоксирибонуклеотидами.
ДНК-полимераза δ
Олигонуклеотид, синтезированный ДНК-полимеразой α и образующий небольшой двухцепочечный фрагмент с матрицей, позволяет присоединиться ДНК-полимеразе 5 и продолжить синтез новой цепи в направлении от 5'- к 3'- концу по ходу раскручивания репликативной вилки.
ДНК-полимераза 5 последовательно наращивает цепь, шаг за шагом присоединяя к ней соответствующие дезоксинуклеотиды. Выбор ДНК-полимеразой 5 очередного нуклеотида определяется матрицей. Включение дезоксирибонуклеозидмонофосфатов в растущую цепь ДНК сопровождается гидролизом макроэргических связей, соответствующих нуклеозидтрифосфатов и отщеплением пирофосфата (Н4Р2О7). Энергия макроэргических связей расходуется на образование 3',5'-фосфодиэфирной связи между последним нуклеотидом растущей цепи ДНК и присоединяемым нуклеотидом. Включение нуклеотида в синтезируемую цепь ДНК невозможно без предварительного связывания азотистого основания водородными связями с комплементарным нуклеотидом матричной цепи. ДНК-полимеразы (α, β, y, δ, ε) могут синтезировать нуклеотидную цепь только в направлении 5' —> 3', матричная цепь всегда считывается в направлении 3' —> 5'.
В каждой репликативной вилке идёт одновременно синтез двух новых (дочерних) цепей. Направление синтеза цепи ДНК совпадает с направлением движения репликативной вилки лишь для одной из вновь синтезируемых цепей (лидирующая цепь). На второй матричной цепи синтез дочерней ДНК осуществляется двумя ферментами: ДНК-полимеразой а и ДНК-полимеразой ε в направлении 5' —> 3', но против движения репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами, которые называют «фрагменты Оказаки» (по имени открывшего их исследователя). Дочерняя цепь ДНК, синтез которой происходит фрагментами, называют отстающей цепью. Каждый фрагмент Оказаки, примерно 100 нуклеотидных остатков, содержит праймер. Праймеры удаляет ДНК-полимераза β, постепенно отщепляя с 5'-конца фрагмента по одному ри- бонуклеотиду. К ОН-группе на 3'-конце предыдущего фрагмента ДНК-полимераза присоединяет дезоксирибонуклеотиды в количестве, равном вырезанному праймеру и таким образом заполняет брешь, возникающую при удалении рибонуклеотидов.
Фермент ДНК-лигаза катализирует образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одного фрагмента цепи ДНК и 5'-фосфатом следующего фрагмента. Реакция протекает с затратой энергии АТФ. Таким образом, из множества фрагментов Оказаки образуется непрерывная цепь ДНК.
В. Ориджины репликации
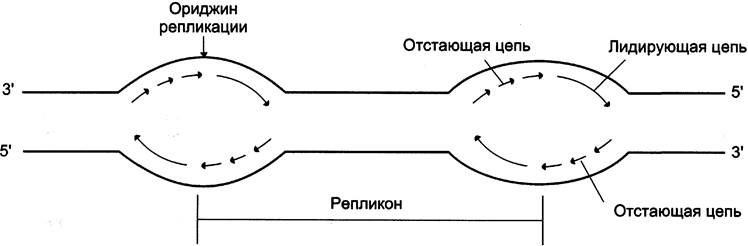
ДНК хромосомы человека содержит примерно 150 млн пар нуклеотидов. Репликация такой большой молекулы со скоростью 50 нуклеотидов в минуту шла бы примерно 800 ч. Поэтому инициация синтеза ДНК происходит в нескольких сайтах хромосомы, которые называют сайтами инициации репликации, или ориджинами (от англ. origin — происхождение) репликации (рис. 4-17). Термин «сайт» используют для обозначения любого участка генома. Ориджины репликации имеют определённую нуклеотидную последовательность. Последовательность ДНК, ограниченную двумя ориджинами репликации, называют единицей репликации, или репликоном. На ориджи- нах при участии ДНК-топоизомеразы I инициируется двунаправленная репликация. Образуются две репликативные вилки, перемещающиеся в противоположных направлениях до тех пор, пока не встретятся со следующим репликоном, т. е. репликация прекращается, когда встречаются две репликативные вилки.
Рис. 4-17. Образование двух репликативных вилок, перемещающихся в противоположных направлениях от ориджина.
Метилирование ДНК
После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей ДНК. Метальные группы присоединяются ко всем остаткам аденина в последовательности -GATC-, при этом образуется N6-метиладенин, а также возможны метилирование цитозина в последовательности -GC- и образование N5-метилцитозина. Количество метилированных оснований равно примерно 1 — 8%. Модификация происходит при участии ферментов, использующих в качестве источника метальных групп S-аденозилметионин (SAM) (см. раздел 9). Присоединение метильных групп к остаткам аденина и цитозина не нарушает комплементарности цепей (рис. 4-18).
Рис. 4-18. Метилирование остатков аденина в последовательности -GАТС-. В течение нескольких минут после репликации, пока не произошло метилирование, новая цепь ДНК отличается от матричной цепи.
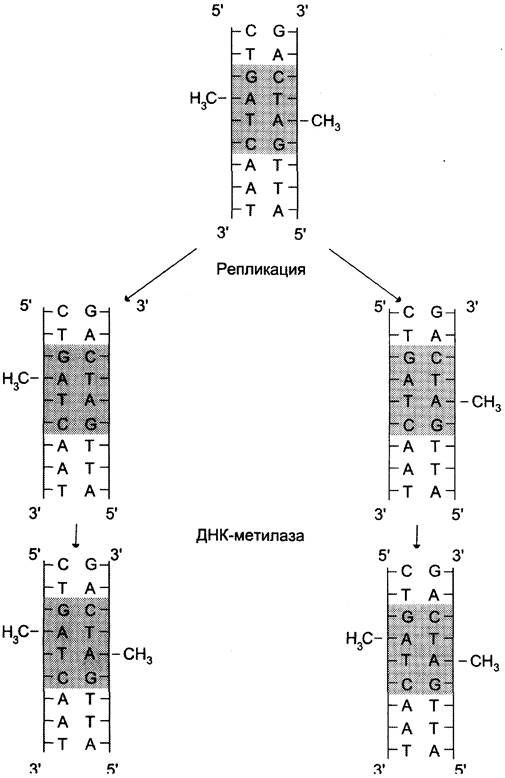
Наличие метальных групп в цепях ДНК необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов. В течение непродолжительного времени в молекуле ДНК последовательности -GATC- метилированы по аденину только в матричной, но не в новой цепи. Это различие используется ферментами репарации для исправления ошибок, которые могут возникать при репликации.
Г. Строение 3' и 5'-концов цепей ДНК. Теломерная ДНК
На каждом конце хромосомы присутствует специфическая нуклеотидная последовательность. Она представлена многочисленными повторами (сотни или даже тысячи раз) олигонуклеотидов -GGGТТА-, называемых теломерной последовательностью, или просто теломерной ДНК. Наличие теломер необходимо для завершения репликации концевых информативных последовательностей хромосом, т. е. для сохранения генетической информации.
После завершения репликации хромосомы 5'-концы дочерних цепей ДНК недостроены, так как после удаления праймеров эти фрагменты оказываются недореплицированными. Это происходит потому, что ДНК-полимераза β, отвечающая за заполнение бреши, образованной после удаления праймера, не может вести синтез цепи ДНК от 3'- к 5'-концу (рис. 4-19, А). Таким образом, в ходе каждого цикла репликации 5'-концы синтезированных цепей укорачиваются. Но такие потери не представляют опасности для генетической информации хромосом, потому что укорочение ДНК идёт за счёт теломер. Во время следующего цикла репликации 5-концы цепей ДНК опять остаются недостроенными. Таким образом, с каждым клеточным делением ДНК хромосом будут последовательно укорачиваться. Укорочение теломер в большинстве клеток по мере их старения — важный фактор, определяющий продолжительность жизни организма.
Рис. 4-19. Синтез теломерной ДНК. А - на рисунке показано укорочение вновь синтезированных цепей ДНК после удаления праймеров; Б - в состав теломеразы входит короткая молекула РНК, содержащая в активном центре последовательность нуклеотидов, комплементарную теломерному повтору; 1 - фермент прикрепляется за счёт взаимодействия РНК с существующей теломерой и добавляет последовательно по одному нуклеотиду фрагмент -GGGТТА-. Матрицей служит простетическая группа теломеразы — фрагмент РНК; 2 - фермент перемещается по нити ДНК таким образом, что РНК-матрица в составе теломеразы постоянно комплементарно связана с концом вновь синтезированного теломерного повтора. Заново синтезированная теломерная ДНК служит матрицей для удлинения второй цепи ДНК, но уже в ходе следующего цикла клеточного деления. Теломерный повтор на рисунке взят в квадратные скобки -[GGGТТА]-.
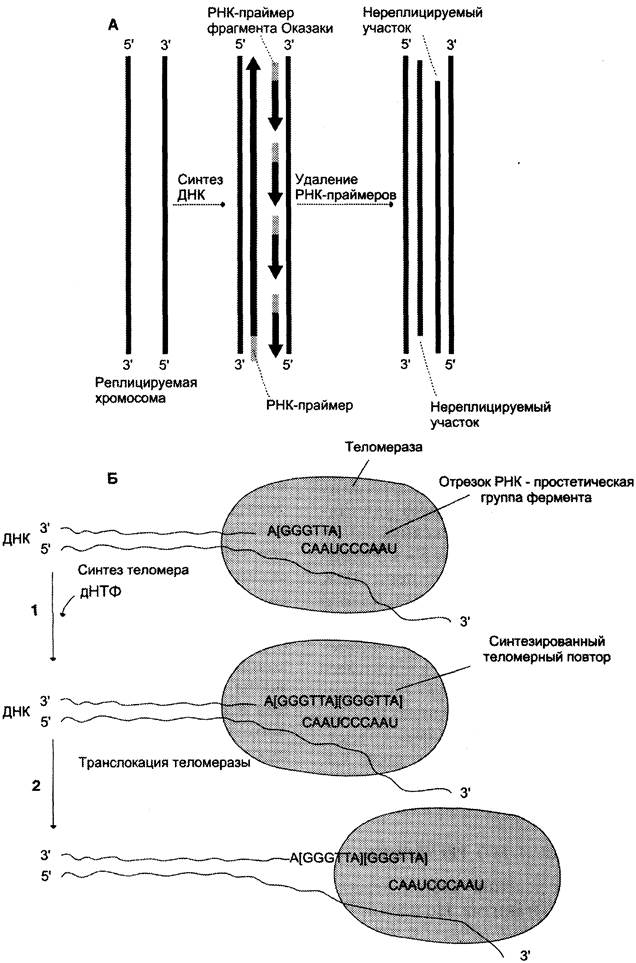
Однако в эмбриональных и других быстро- делящихся клетках потери концов хромосом недопустимы, потому что укорочение ДНК будет происходить очень быстро. В эукариотических клетках имеется фермент теломераза (нуклеотидилтрансфераза), обеспечивающий восстановление недореплицированных 5'-концов. К особенностям этого фермента относят присутствие в качестве простетической группы РНК. Фрагмент РНК в активном центре теломеразы служит матрицей при синтезе теломерных повторов хромосом.
С помощью РНК фермент комплементарно прикрепляется к 3'-концу недостроенной дочерней цепи ДНК. Теломераза по принципу комплементарности последовательно удлиняет 3'-конец цепи ДНК на один гексануклеотид -GGGТТА-. Синтез всегда идёт от 5'- к 3'-концу. Затем теломераза смещается по цепи ДНК на один теломер и начинает синтез нового фрагмента -GGGТТА- (рис. 4-19, Б).
В большинстве соматических клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и её потомства. Однако небольшую активность теломеразы обнаруживают в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия, эпидермиса кожи и др.
Д. Клеточный цикл и его регуляция
Процессы роста и деления клеток лежат в основе жизни любого организма. Но прежде чем совершить деление, клетка должна с высокой точностью копировать свой геном, синтезировать множество высоко- и низкомолекулярных соединений. Совокупность событий, обеспечивающих деление эукариотических клеток, называют «клеточный цикл».Продолжительность клеточного цикла зависит от типа делящихся клеток, у взрослого человека она может варьировать примерно от 8 ч и более, а для некоторых типов клеток до года и больше (рис. 4-20).
Рис. 4-20. Фазы клеточного цикла. После фазы М, в ходе которой происходит деление ядра (митоз) и цитоплазмы (цитокинез), дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы в ходе которой активно происходят биосинтетические процессы, резко замедленные во время митоза. Фаза S — период синтеза ДНК; она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются. Затем наступает фаза G2, в ходе которой происходят деление митохондрий и увеличение энергетических запасов клетки. Фаза G2 продолжается до начала митоза, т. е. фазы М. В фазе М ядерная оболочка разрушается, формируются два новых ядра, цитоплазма делится с образованием двух дочерних клеток, имеющих по одному ядру. На рисунке представлен 24-часовой цикл.
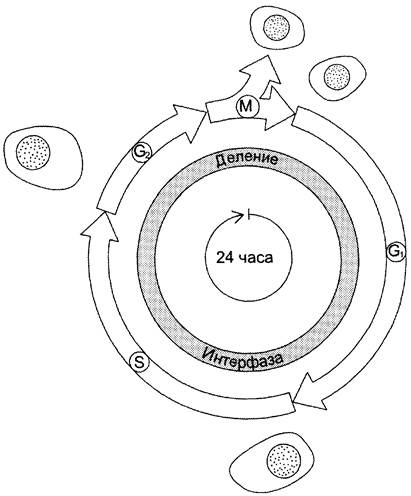
Все фазы клеточного цикла G1, S, G2, М могут различаться по длительности, но в особенности это касается фазы G1, длительность которой может быть равна практически нулю или быть столь продолжительной, что может казаться, будто клетки вообще прекратили деление. В этом случае говорят, что клетки находятся в состоянии покоя (фаза G0). Так, нейроны взрослого человека не делятся вообще. Клетки эпителия кишечника делятся на протяжении всей жизни человека, но даже у этих быстропролиферирующих клеток подготовка к делению занимает 24 ч. Клетки лёгких, почек, печени во взрослом организме начинают делиться только лишь в ответ на повреждение органов.
Внешние сигналы могут стимулировать или ингибировать прохождение клетки через цикл. Пролиферативные сигналы очень разнообразны, они зависят от типа клетки, стадии развития и других факторов. Такими сигналами могут быть факторы роста, интерлейкины, гормоны, способные поддерживать или индуцировать пролиферацию определённых типов клеток. Сигнальные молекулы связываются специфическими мембранными рецепторами, активируют внутриклеточные пути передачи сигналов от рецептора к ядру и таким образом индуцируют транскрипцию определённых генов. Одними из первых активируются гены, кодирующие белки циклины. Белки были названы циклинами, потому что их концентрация в клетке периодически меняется по мере прохождения клеткой разных фаз клеточного цикла.
Все циклины делят на 2 подсемейства: G1- циклины (D, Е) и митотические циклины (А и В). Любой из циклинов представлен группой полиморфных белков, например, циклин Dпредставлен формами D1, D2, D3. У каждого типа циклинов есть гомологичный участок из 100 аминокислотных остатков — «циклиновый бокс», отвечающий за связывание с циклинзависимой киназой (от англ. CDK — cyclin- dependent kinases). В клетках эукариотов существует примерно восемь различных CDK (CDK1-8), активирующихся различными циклинами (табл. 4-2).
Таблица 4-2. Циклины и циклинзависимые киназы, регулирующие прохождение клеточного цикла
Циклин |
Киназа |
Функция |
D, Е |
CDK4, CDK6 |
Регулирует переход клетки из в G1 -фазы в S-фазу |
А |
CDK2 I |
Активирует синтез ДНК на начальной стадии S-фазы |
В |
CDK1 |
Регулирует переход клетки из G2-фазы в М-фазу |
Циклинзависимые киназы, связывая циклин, переходят в активную форму и могут фосфорилировать специфические белки, например, факторы транскрипции, белки-ингибиторы факторов транскрипции, которые регулируют синтез ферментов, обеспечивающих репликацию. Синтез каждого циклина начинается при подготовке к соответствующей фазе клеточного цикла, его концентрация в клетке повышается, а после окончания фазы резко падает до нуля. Завершившие свою работу комплексы циклинов и CDK связываются специфическими белками, ингибирующими их активность, и затем подвергаются разрушению.