БИОХИМИЯ УЧЕБНИК ДЛЯ ВУЗОВ - Е. С. Северина - 2004
РАЗДЕЛ 11. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВЕЩЕСТВ И ФУНКЦИЙ ОРГАНИЗМА
VIII. Роль гормонов в регуляции репродуктивной функции организма
Репродуктивные функции организма регулируются половыми гормонами: у мужчин — тестостероном, у женщин — эстрогенами и прогестинами. Синтез и секреция половых гормонов, в свою очередь, находятся под контролем фолликулостимулирующего и лютеинизирующего гормонов.
А. Гонадотропные гормоны гипофиза, стимулирующие синтез и секрецию половых гормонов
Фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ) — гонадотропные гормоны гипофиза. Представляют собой гликопротеины с молекулярной массой около 30 кД, состоящие из α- и β-субъединиц. α-Субъединицы содержат 92 аминокислоты и две боковые углеводные цепи и идентичны α-субъединице тиреотропина. β-Субъединицы индивидуальны для каждого гормона.
1. Регуляция секреции ФСГ и ЛГ
Образование и освобождение обоих гормонов стимулируется гипоталамическим декапептидом — гонадотропин-рилизинг-гормоном, секреция которого происходит эпизодически, что в основном и определяет импульсный характер секреции ЛГ и ФСГ.
У женщин эстрогены и прогестерон по механизму обратной связи влияют на секрецию ЛГ и ФСГ как на гипоталамическом, так и на гипофизарном уровне.
У мужчин тестостерон и эстроген, образованный в клетках Лейдига и в процессе метаболизма тестостерона, блокируют по механизму обратной связи синтез и секрецию гонадолиберина и гонадотропных гормонов гипофиза. Кроме этого, клетками гранулёзы фолликулов и клетками Сертоли вырабатывается белок ингибин, который тормозит гипофизарную секрецию ФСГ.
Т1/2 ФСГ составляет примерно 150 мин, а Т1/2 ЛГ — 30 мин.
2. Механизм действия и эффекты ФСГ и ЛГ
Гонадотропные гормоны ЛГ и ФСГ связываются с рецепторами на мембранах своих клеток-мишеней в яичниках и яичках, в результате чего происходит активация аденилатциклазной системы. Образующийся цАМФ активирует протеинкиназу, которая фосфорилирует белки, опосредующие эффекты ЛГ и ФСГ.
У женщин лютеинизирующий гормон стимулирует образование прогестерона клетками жёлтого тела, у мужчин — синтез тестостерона интерстициальными клетками Лейдига. ФСГ ускоряет развитие фолликулов в яичниках и образование эстрогенов, а действуя на клетки Сертоли, запускает процесс сперматогенеза.
Б. Мужские половые гормоны
Мужские половые гормоны (рис. 11-39) вырабатываются в основном в мужских половых железах — в интерстициальных клетках Лейдига семенников (95%). Небольшое количество андрогенов образуется в коре надпочечников.
Рис. 11-39. Мужские половые гормоны.
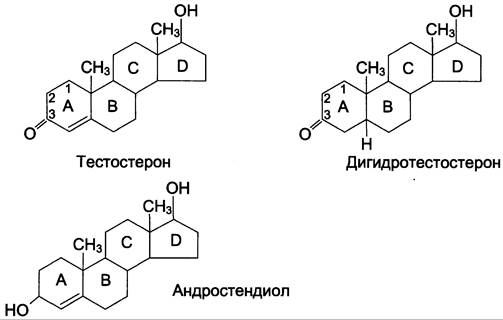
1. Синтез андрогенов
Путь биосинтеза андрогенов в яичках и коре надпочечников одинаков. Предшественником андрогенов, как и других стероидных гормонов, служит холестерол (рис. 11-40), который либо поступает из плазмы в составе ЛПНП, либо синтезируется в самих железах из ацетил-КоА.
Рис. 11-40. Схема синтеза половых гормонов. Предшественником половых гормонов служит холестерол. Образование прегненолона происходит в результате отщепления боковой цепи холестерола (1). Превращение прегненолона в тестостерон может протекать двумя путями: через образование прогестерона (2) или дегидроэпиандростерона (3). Тестостерон служит предшественником дигидротестостерона (4). В некоторых периферических тканях небольшое количество тестостерона превращается в эстрадиол (5). В яичниках синтезируются женские половые гормоны, эстрогены и прогестины, среди которых наиболее активными являются 17β-эстрадиол и прогестерон. Ароматизация андрогенов протекает под действием ароматазного комплекса, содержащего цитохром Р450-оксидазу, и включает 3 реакции гидроксилирования с участием O2 и NАDРH.
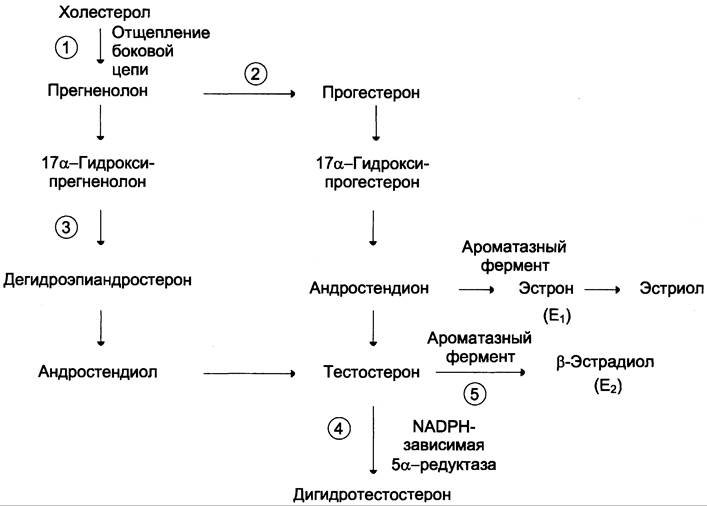
Отщепление боковой цепи холестерола и образование прегненолона — скорость — лимитирующая реакция. Однако, в отличие от аналогичной реакции, протекающей в надпочечниках, эта стадия стимулируется ЛГ (а не АКТГ). ЛГ, связываясь с рецептором плазматической мембраны клеток Лейдига, активирует аденилатциклазу, увеличивая тем самым внутриклеточную концентрацию цАМФ, что в конечном итоге вызывает активацию фермента, который расщепляет боковую цепь холестерола между С-20 и С-22.
Тестостерон. Превращение прегненолона в тестостерон катализируется пятью микросомальными ферментами и может протекать двумя путями: через образование дегидроэпиандростерона или через образование прогестерона (что, по-видимому, преобладает в семенниках человека).
Суточная секреция тестостерона у мужчин составляет в норме примерно 5 мг и сохраняется на протяжении всей жизни организма. Гормон циркулирует в крови в связанном с белками плазмы состоянии: альбумином (40%) и специфически связывающим половые гормоны β-глобулином (называемым секс-гормонсвязывающим глобулином, СГСГ). Лишь 2% от общего количества гормона в крови транспортируется в свободном виде, и именно такие молекулы проявляют биологическую активность.
Дигидротестостерон. В семенных канальцах, предстательной железе, коже, наружных половых органах тестостерон служит предшественником более активного андрогена — дигидротестостерона (рис. 11-41, 11-42). Это превращение, в котором участвует примерно 4% тестостерона, происходит в результате восстановления двойной связи кольца А и 3-кето- группы при участии цитоплазматического фермента — NАDРН-зависимой 5α-редуктазы. Семенники человека секретируют в сутки до 50 — 100 мкг дигидротестостерона. Однако большее количество гормона — следствие периферических превращений, и суммарная суточная секреция дигидротестостерона составляет 400 мкг, что почти в 10 раз меньше уровня секреции тестостерона.
В некоторых периферических тканях небольшое количество тестостерона превращается в эстрадиол. В качестве побочных продуктов клетки Лейдига также постоянно секретируют эстрадиол и прогестерон, хотя роль этих гормонов в развитии и поддержании функций размножения и формирования полового поведения у мужчин до настоящего времени не выяснена.
2. Регуляция синтеза и секреции андрогенов
В препубертатный период секреция андрогенов подавляет по механизму отрицательной обратной связи секрецию гонадотропина до начала пубертатного периода, когда гипофизарные клетки становятся менее чувствительными к ингибирующему действию циркулирующих в крови андрогенов. Эта потеря чувствительности приводит к циклически импульсному освобождению ЛГ и ФСГ. ЛГ, связываясь с рецепторами клеток Лейдига, стимулирует образование тестостерона интерстициальными клетками Лейдига, а ФСГ, связываясь с рецепторами клеток Сертоли в семенниках, стимулирует сперматогенез (рис. 11-41).
Рис. 11-41. Регуляция синтеза и секреции мужских половых гормонов. Синтез и секреция мужских половых гормонов регулируется гипоталамо-гипофизарной системой по механизму отрицательной обратной связи. Секреция ЛГ и ФСГ стимулируется гонадотропин-рилизинг гормоном. ЛГ ускоряет синтез и секрецию тестостерона клетками Лейдига, ФСГ стимулирует сперматогенез. Тестостерон стимулирует сперматогенез, ингибирует синтез и секрецию гонадотропин-рилизинг гормона и ЛГ.
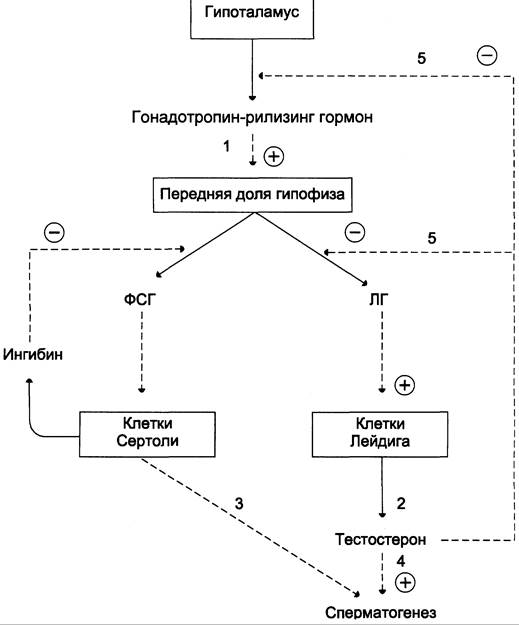
Тестостерон замыкает отрицательную обратную связь на уровне гипофиза и гипоталамуса, уменьшая частоту секреторных импульсов ЛГ. Торможение секреции ФСГ аденогипофизом происходит под действием белка ингибина, вырабатываемого клетками Сертоли. ФСГ стимулирует синтез этого белка, который по механизму отрицательной обратной связи тормозит дальнейшую секрецию ФСГ.
3. Мишени для андрогенов
К мишеням тестостерона относят эмбриональные вольфовы структуры, сперматогонии, мышцы, кости, почки, мозг. Подобно другим стероидным гормонам, андрогены образуют внутри клетки комплекс с рецептором, который связывается с определённым участком хроматина, активируя специфические гены, белковые продукты которых опосредуют биологические эффекты андрогенов.
4. Эффекты андрогенов
Физиологическое действие андрогенов различно в разные периоды жизни организма. У эмбриона под действием андрогенов из вольфова протока образуются придаток яичка (эпидидимис), семявыносящий проток и семенной пузырёк. У плода мужского пола происходит маскулинизация мозга. Поскольку андрогены в организме обладают мощным анаболическим действием и стимулируют клеточное деление, повышенный уровень андрогенов в препубертатный период приводит к скачкообразному увеличению линейных размеров тела, увеличению скелетных мышц, росту костей, но одновременно способствуют и остановке роста, так как стимулируют сращение эпифизов длинных костей с их стволами. Андрогены вызывают изменение структуры кожи и волос, снижение тембра голоса вследствие утолщения голосовых связок и увеличения объёма гортани, стимулируют секрецию сальных желёз.
В. Женские половые гормоны
В яичниках синтезируются женские половые гормоны — эстрогены и прогестины, среди которых наиболее активны 17β-эстрадиол и прогестерон (рис. 11-42).
Рис. 11-42. Женские половые гормоны.
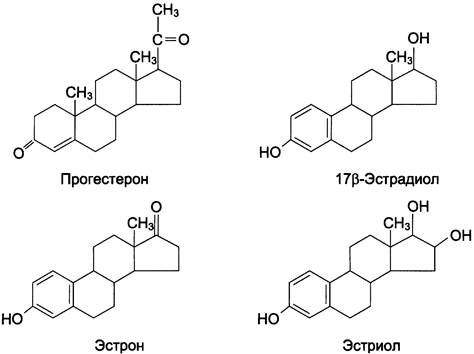
1. Образование эстрогенов
Согласно современным представлениям, образование эстрогенов яичников предполагает выработку андрогенов (андростендиона) в клетках теки фолликулов с последующей ароматизацией андрогенов в клетках гранулёзы. В клетках теки синтезируются рецепторы ЛГ. Рецепторы ФСГ образуются в клетках гранулёзы. ЛГ, связываясь с рецепторами клеток теки и активируя фермент, который катализирует отщепление боковой цепи холестерола и превращение его в прегненолон, тем самым стимулирует образование основного андрогена яичников — андростендиона. ФСГ, взаимодействуя с рецепторами клеток гранулёзы, активирует содержащийся в этих клетках ароматазный ферментативный комплекс и стимулирует превращение андрогенов, вырабатываемых клетками теки, в эстрогены. Ароматизация андрогенов под действием ароматазного комплекса, содержащего цитохром Р450-оксидазу, включает 3 реакции гидроксилирования, которые протекают с участием О2 и NADPH.
Непосредственно в клетках теки синтезируется очень небольшое количество эстрогенов. Значительная часть эстрогенов продуцируется путём периферической ароматизации андрогенов в жёлтом теле, фетоплацентарном комплексе (во время беременности), корой надпочечников, в жировых клетках, печени, коже и других тканях, где обнаружена повышенная ароматазная активность.
В клетках гранулёзы может синтезироваться менее активный эстроген — эстрон, а ещё менее активный эстриол образуется из эстрона в крови. В печени β-эстрадиол инактивируется в результате гидроксилирования ароматического кольца по атому углерода С2 и образования конъюгатов с серной или глюкуроновой кислотами, которые и выводятся из организма с жёлчью или мочой.
Примерно 95% циркулирующих в крови эстрогенов связано с транспортными белками — СГСБ (секс-гормонсвязывающий белок) и альбумином. Биологической активностью обладает только свободная форма эстрогенов.
2. Регуляция секреции эстрогенов
В детском возрасте незрелые яичники вырабатывают небольшое количество гормонов, поэтому концентрация эстрогенов в крови низкая. В пубертатный период чувствительность гипоталамо-гипофизарной системы к действию ЛГ и ФСГ снижается. Импульсная секреция гонадотропин-рилизинг-гормона устанавливает суточный ритм секреции ЛГ и ФСГ. В начале каждого менструального цикла секреция ФСГ и ЛГ вызывает развитие первичных фолликулов. Созревающий фолликул в результате совместного действия ЛГ, стимулирующего продукцию андрогенов клетками теки, и ФСГ, стимулирующего ароматизацию андрогенов, секретирует эстрогены, которые по механизму отрицательной обратной связи угнетают секрецию ФСГ. Концентрация ФСГ в крови остаётся низкой ещё и в результате торможения секреции этого гормона белком ингибином, выделяемым яичниками (рис. 11-43).
Рис. 11-43. Регуляция секреции женских половых гормонов. Гонадотропин-рилизинг гормон стимулирует секрецию ЛГ и ФСГ, которые совместно с эстрогеном и прогестероном регулируют половой цикл у женщин. Эстрадиол и прогестерон по механизму отрицательной обратной связи регулируют синтез и секрецию ЛГ и ФСГ.
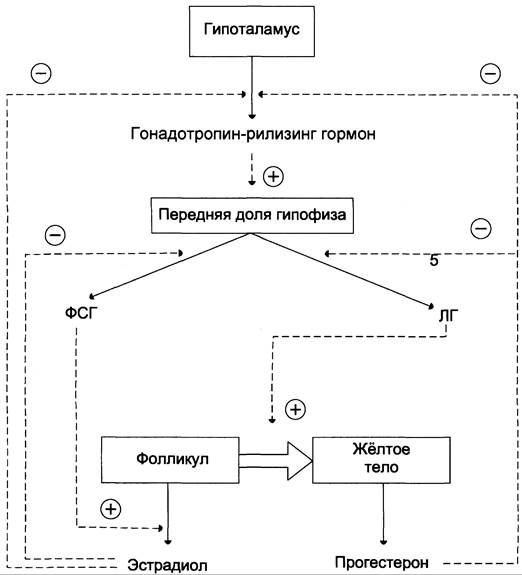
По мере созревания фолликула (фолликулярная фаза) концентрация эстрадиола повышается, чувствительность гипофизарных клеток к гонадолиберину возрастает, и эстрадиол по механизму положительной обратной связи повышает секрецию ЛГ и ФСГ.
Повышение секреции ЛГ приводит к овуляции — освобождению яйцеклетки из лопнувшего фолликула. После овуляции клетки гранулёзы превращаются в жёлтое тело, которое, помимо эстрадиола, начинает вырабатывать всё большее количество основного гормона лютеиновой фазы — прогестерона (прогестина). Если возникает беременность, жёлтое тело продолжает функционировать и секретировать прогестерон, однако на более поздних этапах беременности прогестерон в основном продуцируется плацентой. Если оплодотворение не происходит, высокая концентрация прогестерона в плазме крови по механизму отрицательной обратной связи угнетает активность гипоталамо-гипофизарной системы, тормозится секреция ЛГ и ФСГ, жёлтое тело разрушается, и снижается продукция стероидов яичниками. Наступает менструация, которая длится примерно 5 дней, после чего начинает формироваться новый поверхностный слой эндометрия, и возникает новый цикл.
3. Механизм действия и биологические эффекты эстрогенов
Эстрогены связываются с внутриклеточными рецепторами и, подобно другим стероидным гормонам, регулируют транскрипцию структурных генов. Предполагается, что эстрогены индуцируют синтез свыше 50 различных белков, участвующих в проявлении физиологических эффектов эстрогенов.
Эстрогены стимулируют развитие тканей, участвующих в размножении, определяют развитие многих женских вторичных половых признаков, регулируют транскрипцию гена рецептора прогестина. В лютеиновой фазе под действием эстрогенов вместе с прогестинами пролиферативный эндометрий (эпителий матки) превращается в секреторный, подготавливая его к имплантации оплодотворённой яйцеклетки. Совместно с простагландином F2α эстрогены увеличивают чувствительность миометрия к действию окситоцина во время родов. Эстрогены оказывают анаболическое действие на кости и хрящи. Другие метаболические эффекты эстрогенов включают поддержание нормальной структуры кожи и кровеносных сосудов у женщин, способствуют образованию оксида азота в сосудах гладких мышц, что вызывает их расширение и усиливает теплоотдачу. Эстрогены стимулируют синтез транспортных белков тиреоидных и половых гормонов. Эстрогены могут индуцировать синтез факторов свёртывания крови II, VII, IX и X, уменьшать концентрацию антитромбина III.
Эстрогены оказывают влияние на обмен липидов. Так, увеличение скорости синтеза ЛПВП и торможение образования ЛПНП, вызываемое эстрогенами, приводит к снижению содержания холестерола в крови.
4. Образование прогестерона
Прогестерон, образующийся главным образом жёлтым телом во время менструации в лютеиновую фазу, секретируется также фетоплацентарным комплексом во время беременности. В небольших количествах он вырабатывается у женщин и мужчин корой надпочечников. В фолликулярной фазе менструального цикла концентрация прогестерона в плазме обычно не превышает 5 нмоль/л, а в лютеиновой фазе увеличивается до 40-50 нмоль/л. В крови прогестерон связывается с транспортным глобулином транскортином и альбумином, и только 2% гормона находится в свободной биологически активной форме. Диффундируя в клетки-мишени, прогестерон связывается со специфическим ядерным рецептором. Образующийся комплекс гормон — рецептор взаимодействует с промоторным участком ДНК и активирует транскрипцию генов. Т1/2прогестерона в крови составляет 5 мин. В печени гормон конъюгируется с глюкуроновой кислотой и выводится с мочой.
5. Биологические эффекты прогестерона
Действие прогестерона в основном направлено на репродуктивную функцию организма. Образование прогестерона отвечает за увеличение базальной температуры тела на 0,2 — 0,5 °С, которое происходит сразу после овуляции и сохраняется на протяжении лютеиновой фазы менструального цикла. При высоких концентрациях прогестерон взаимодействует с рецепторами, локализованными в клетках почечных канальцев, конкурируя таким образом с альдостероном. В результате конкурентного игибирования альдостерон теряет возможность стимулировать реабсорбцию натрия.
Прогестерон может также оказывать действие и на ЦНС, в частности вызывать некоторые особенности поведения в предменструальный период.