Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ VI. БІОХІМІЯ ФІЗІОЛОГІЧНИХ ФУНКЦІЙ ТА СПЕЦІАЛІЗОВАНИХ ТКАНИН
ГЛАВА 26. БІОХІМІЯ ХАРЧУВАННЯ ЛЮДИНИ. І. КОМПОНЕНТИ ХАРЧУВАННЯ. ТРАВЛЕННЯ ПОЖИВНИХ РЕЧОВИН
26.3. МЕХАНІЗМИ ПЕРЕТВОРЕННЯ ПОЖИВНИХ РЕЧОВИН У ТРАВНОМУ ТРАКТІ
Перетравлення поживних речовин (білків, вуглеводів, ліпідів) — це процес гідролізу відповідних сполук у складі продуктів харчування, що відбувається в травному каналі і призводить до утворення простих біомолекул, які за рахунок дії спеціальних механізмів мембранного транспорту всмоктуються у кров.
Початкові процеси травлення відбуваються в ротовій порожнині за участю слини, яка є біологічною рідиною з рН 6,8, що на 99,5 % складається з води і містить різноманітні білки (ферменти, муцини, імуноглобуліни, лізоцим тощо) та неорганічні солі. Слина має змащувальну дію відносно сухих продуктів харчування, під її впливом полегшується процес жування та створюються умови для подальшого перетворення компонентів харчування під впливом специфічних ферментів.
До ферментів слини належать глікозидази, що каталізують певні процеси гідролізу вуглеводів — α-амілаза та мальтаза. Під дією цих ферментів можливе розщеплення крохмалю до високомолекулярних декстринів і мальтози до глюкози, але, оскільки час впливу слини на харчову грудку незначний, — відповідні продукти утворюються в порожнині рота лише в незначній кількості.
Основні процеси травлення поживних речовин їжі відбуваються в шлунку — розщеплення білків до пептидних молекул та в різних відділах тонкого кишечника — розщеплення пептидів, вуглеводів, жирів (ліпідів). Шлунок виробляє власні протеолітичні ферменти (пепсиноген, ренін); кишечник (залози Брунера та Ліберкюна) синтезує деякі пептидази, дисахаридази, фосфоліпази та полінуклеотидази. Травлення в кишечнику неможливе без участі гідролітичних ферментів, які надходять сюди з підшлункової залози — протеаз (трипсину, хімотрипсину,
еластази), карбоксипептидази, амілаз, ліпаз. У процесі травлення жирів беруть участь біохімічні компоненти жовчі, що синтезуються в гепатоцитах печінки.
Перетравлення білків
Біохімічні процеси перетравлення білків та пептидів, що надходять до організму людини з їжею, відбуваються в шлунку й тонкій кишці. Гідроліз цих компонентів відбувається під дією ферментів, які виробляються клітинами слизової оболонки травного каналу й екзокринної частини підшлункової залози. Протеази шлунка, кишечника та підшлункової залози гідролізують певні пептидні зв’язки в молекулах білків і пептидів їжі, і в результаті їх послідовної дії утворюється суміш вільних L-амінокислот та найпростіших пептидів, що транспортуються всередину ентероцитів і далі — в кров’яне русло.
Перетравлення білків у порожнині шлунка
Шлунковий сік, під дією якого відбувається гідроліз білків, — це кисла рідина з рН 1,5-2,5. Основними біохімічними компонентами шлункового соку, що беруть участь в перетворенні білків продуктів харчування, є соляна кислота та протеолітичний фермент пепсин. Крім того, до складу шлункового соку входять кислі фосфати (переважно NaH2PO4) та деякі органічні кислоти, складаючи загальну кислотність шлунка.
Соляна кислота виробляється в спеціальних обкладинних (оксинтних) клітинах слизової оболонки шлунка за участю хлоридів, які надходять із крові. Донором протонів, необхідних для утворення HCl, є вугільна кислота, що утворюється з Н2О та СО2 за участю карбоангідрази:
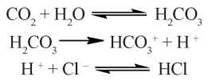
Секреція іонів Н+ в порожнину шлунка відбувається при дії протонної помпи мембран оксинтних клітин — Н+, К+-АТФази (рис. 26.2).
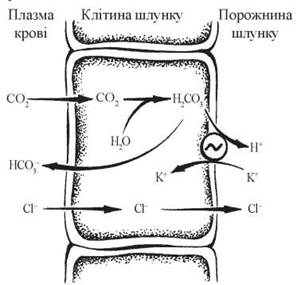
Рис. 26.2. Схема утворення соляної кислоти в шлунку, ~ Н+, К+-АТФаза.
Концентрація HCl у шлунковому соку складає 0,45-0,60 %. Соляна кислота необхідна для утворення активного ферменту пепсину і прояву максимуму його каталітичної активності.
Пепсин — протеаза з м.м. 42 кД, що синтезується головними клітинами слизової оболонки шлунка у вигляді проферменту пепсиногену (м.м. 35 кД). Початкові етапи перетворення пепсиногену в пепсин здійснюються за участю іонів Н+, які сприяють відщепленню від молекули проферменту N-кінцевого захисного пептиду, що супроводжується розкриттям активного центру; в подальшому процес стає авто- каталітичним — молекули пепсину спричиняють власне утворення з проферменту:
![]()
За механізмом дії пепсин є ендопептидазою, що специфічно атакує пептидні зв’язки, в утворенні яких беруть участь залишки ароматичних (фенілаланіну, тирозину), а також дикарбонових (глутамату, аспартату) амінокислот. Під дією пепсину білки розшеплюються на великі поліпептидні фрагменти — пептони, гідроліз яких завершується в тонкій кишці.
Реннін (хімозин, сичужний фермент) — протеаза, що міститься в шлунковому соку новонароджених дітей. Реннін є ферментом, який за участю іонів Са2+ спричиняє перетворення розчинних білків молока — казеїнів у нерозчинні — параказеїни, які підлягають протеолітичній дії пепсину («створожування молока»).
Перетравлення білків в кишечнику
Частково перетравлена напіврідка маса поживних сполук, що утворюється в шлунку (хімус) періодично надходить через пілоричний клапан у дванадцятипалу кишку. В цю ж частину травного каналу надходять із підшлункової залози протеолітичні ферменти та пептидази, які діють на пептиди, що надходять зі шлунка. Каталітична дія цих ферментів відбувається в слабколужному середовищі (рН 7,5-8,0), яке утворюється наявними в кишковому соку бікарбонатами NaHCO3.
Більшість ферментів протеолітичної дії, що функціонують у тонкій кишці, синтезуються в екзокринних клітинах підшлункової залози у вигляді проферментів, які активуються після їх надходження в дванадцятипалу кишку (трипсиноген, хімотрипсиноген, проеластаза, прокарбоксипептидази А і В).
Гідроліз білків та пептидів, що надходять із шлунка, відбувається як у порожнині тонкої кишки, так і на поверхні ентероцитів — пристінкове, або мембранне травлення.
Трипсин — протеолітичний фермент з м.м. 24,7 кД, що утворюється в порожнині кишечника з неактивного проферменту трипсиногену під дією ентерокінази, яка відщеплює від молекули проферменту N-кінцевий гексапептид з утворенням каталітично активного трипсину:
![]()
Трипсин є ендопептидазою, яка найбільш активна відносно пептидних зв’язків, утворених основними амінокислотами аргініном та лізином.
Хімотрипсин — протеолітичний фермент (м.м. 29 кД), який утворюється з проферменту хімотрипсиногену за каталітичної дії трипсину, що відщеплює від молекули проферменту декілька інгібіторних пептидів:
![]()
Хімотрипсин є ендопептидазою, яка розщеплює до 50 % пептидних зв’язків в молекулах білків та пептидів їжі, в тому числі зв’язків, нечутливих до дії пепсину та трипсину.
Еластаза — ендопептидаза, що також має широку субстратну специфічність, розщеплюючи пептидні за’язки, що утворюються залишками амінокислот малого розміру — гліцину, аланіну, серину.
Утворені при дії зазначених вище ендопептидаз короткі пептиди підлягають дії екзопептидаз кишечника — карбоксипептидаз А і В, амінопептидаз та дипептидаз.
Карбоксипептидази — пептидази, що гідролізують пептидні зв’язки, утворені С-кінцевими амінокислотами: карбоксипептидаза А відщеплює від С-кінця амінокислоти з гідрофобними радикалами, а карбоксипептидаза В — С-кінцеві залишки лізину й аргініну.
Амінопептидази — ферменти ентероцитів, що відщеплюють від коротких пептидів N-кінцеві амінокислотні залишки.
Дипептидази — пептидогідролази, що розщеплюють дипептиди до вільних амінокислот.
Послідовна дія всього набору шлункових, панкреатичних і кишечних пептидогідролаз забезпечує повне розщеплення білків та пептидів продуктів харчування до амінокислот. У кровотік слизовою оболонкою кишечника всмоктуються тільки вільні амінокислоти.
Перетравлення вуглеводів
Основні реакції розщеплення вуглеводів відбуваються в тонкому кишечнику за рахунок дії ферментів підшлункової залози, що потрапляють у порожнину дванадцятипалої кишки, і власних ферментів кишкового соку. Подібно до перетворення білків та пептидів, поряд з порожнинним травленням, у кишечнику відбувається пристінкове (на поверхні мембран ентероцитів) травлення вуглеводів.
Амілази, що діють у кишечнику — це ферменти α-амілаза (переважно) та β-амілаза, які синтезуються в підшлунковій залозі. Панкреатична а-амілаза — це ендоглікозидаза, подібна до ферменту слини, яка гідролізує крохмаль та глікоген з утворенням суміші розгалужених і нерозгалужених олігосахаридів і деякої кількості мальтози і мальтотріози. β-Амілаза — панкреатична екзоглікозидаза, яка відщеплює від нерозгалужених гомополісахаридних ланцюгів залишки мальтози. Гідроліз гомополісахиридів у точках розгалуження (1→б) каталізується α(1→б)- глікозидазою.
Дисахаридази та олігосахаридази — ферменти, що синтезуються в тонкій кишці і спричиняють розщеплення до моносахаридів відповідних цукрів, які утворюються як продукти дії амілаз або надходять до травного каналу в складі рослинних продуктів харчування:
мальтаза (а-глюкозидаза) — фермент, що гідролізує мальтозу та відщеплює термінальні глюкозні залишки з нередукуючих кінців α(1→4)-зв’язаних олігосахаридів; мальтаза та ізомальтаза(α(1→6)-глікозидаза) завершують розщеплення гомополісахаридів, розпочате амілазами;
лактаза (β-галактозидаза) — фермент, що розщеплює лактозу (молочний цукор) до двох моносахаридів — галактози та глюкози; надзвичайно велике фізіологічне значення лактази в харчуванні дітей;
сахараза (β-фруктозидаза) — фермент кишечного соку, що гідролізує з утворенням глюкози і фруктози дисахарид сахарозу — основний компонент бурякового та тростинного цукру.
Внаслідок дії зазначених глікозидазних ферментів на рослинні та тваринні вуглеводи продуктів харчування утворюється суміш моносахаридів (в основному глюкози, фруктози й галактози), які всмоктуються клітинами кишкового епітелію і поступають у кров. Глюкоза складає до 90 % усіх моносахаридів крові, решту становлять інші гексози та пентози, утворюючи в сумі загальний цукор крові (4,5-6,5 ммоль/л).
Недостатність дисахаридаз
Існує група спадкових ензимопатій, що пов’язані з недостатністю синтезу і виділення в кишковий сік ферментів, які гідролізують дисахариди. Ці ферментні дефекти проявляються порушеннями у перетравленні та всмоктуванні відповідних цукрів.
Недостатність лактази
Спадковий дефіцит ферменту призводить до неспроможності кишкового соку розщеплювати молочний цукор, і позначається як непереносимість лактози.
У відносно рідкісних випадках патологія (спадкова відсутність лактази) в умовах харчування материнським молоком клінічно проявляється вже в перші дні життя новонародженого. Проте, в більшості випадків ензимопатія зустрічається у вигляді низької активності лактази. Ця форма непереносимості лактози успадковується як автосомна рецесивна патологія і вперше проявляється в підлітковому періоді або у молодому віці. Розповсюдженість недостатності лактази широко варіює в різних расових групах, особливо часто зустрічаючись у жителів східних країн та кольорового населення Північної Америки. Так, зокрема частота патології у датчан становить 3 %, а в тайців — 97 %.
Недостатність сахарази звичайно виявляється разом із недостатністю ізомальтази у вигляді сполученого дефекту — непереносимості двох дисахаридів. Ферментопатія проявляється після переводу новонароджених на мішане харчування з додаванням фруктових соків та інших продуктів, що містять рослинні цукри.
Клінічно недостатність дисахаридаз проявляється симптомами вуглеводної диспепсії — діареєю, метеоризмом; новонароджені діти відстають у розвитку.
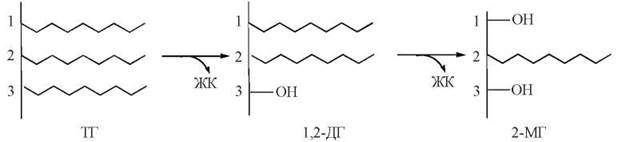
Рис. 26.3. Переварювання нейтральних жирів під дією панкреатичної ліпази: ТГ - триацилгліцерол; 1,2-ДГ - 1,2-діацилгліцерол; 2-МГ - 2-моноацилгліцерол; ЖК - жирна кислота.
Перетравлення ліпідів
Перетравлення ліпідів харчових продуктів здійснюється в дванадцятипалій кишці під впливом ферментів, що синтезуються в неактивній формі в екзокринних клітинах підшлункової залози, а саме — ліпази, фосфоліпази А2, холестеролестерази та власних ферментів кишечника.
1. Гідроліз нейтральних жирів відбувається за рахунок дії ліпази підшлункової залози. Панкреатична ліпаза специфічна до складноефірних зв’язків в положеннях 1 та 3 триацилгліцеролів, тому продуктами дії ферменту є вільні жирні кислоти та 2-моноацилгліцерол:
Активне функціонування панкреатичної ліпази реалізується за умов оптимальної лужності (рН панкреатичного секрету = 7,5-8,0) та наявності амфіпатичних молекул жовчних кислот (головним чином — глікохолевої і таурохолевої), які необхідні для емульгування харчових жирів і утворення міцел триацилгліцеролів. Процес взаємодії ферментного білка ліпази з поверхнею розподілу фаз у системі жовчно-кисла сіль/триацилгліцерол потребує також наявності додаткового фактора — білка коліпази, який міститься в секреті підшлункової залози.
Крім емульгування харчових жирів, що є передумовою дії панкреатичної ліпази, жовчні кислоти беруть також участь у формуванні і всмоктуванні міцелярних структур (вільні жирні кислоти, моногліцериди), що формуються після гідролізу триацилгліцеролів.
Основна маса жовчних кислот (9095 % їх загальної кількості)
всмоктується з кров’ю v. porta в нижніх відділах тонкї кишки і надходить у печінку, де повторно використовується для формування жовчі — процес ентерогепатичної циркуляції. Таким чином, з каловими масами щодобово виводиться лише до 0,5 г жовчних кислот; їх втрата компенсується за рахунок синтезу в гепатоцитах нових молекул первинних жовчних кислот (холевої та хенодезоксихолевої) з холестерину.
2. Гідроліз фосфоліпідів (гліцерофосфоліпідів) каталізується фосфоліпазою А2, яка синтезується в підшлунковій залозі у вигляді проферменту та перетворюється в активну форму шляхом триптичного гідролізу певних пептидних зв’язків у молекулі каталітично неактивного білка.
Панкреатична фосфоліпаза А2 гідролізує складноефірні зв’язки в положенні 2 фосфогліцеридів з утворенням лізофосфоліпідів. Інші фосфоліпази, що містяться в кишковому соку, розщеплюють гліцерофосфоліпіди до гліцерину, вищих жирних кислот, азотистих основ та фосфорної кислоти:
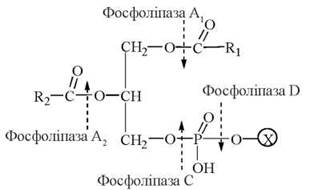
Рис. 26.4. Схема гідролізу молекули гліцерофосфоліпіду (Х - азотиста основа).
3. Гідроліз ефірів холестерину відбувається під дією холестеролестерази (гідролази холестерилових ефірів) з утворенням холестеролу, який всмоктується ентероцитами у вільній формі.
У результаті розглянутих біохімічних процесів, що відбуваються з харчовими ліпідами в порожнині кишечника, утворюється складна суміш продуктів, основними компонентами якої є:
- вільні вищі жирні кислоти (у вигляді Na+ та К+-солей);
- 2-моноацилгліцероли;
- вільний (неетерифікований) холестерин;
- продукти гідролізу гліцерофосфоліпідів (гліцерин, аміноспирти, солі фосфорної кислоти);
- триацилгліцероли, що містять залишки коротколанцюгових (переважно С8-С10) жирних кислот. Такі триацилгліцероли складають до 10 % від загальної кількості нейтральних жирів їжі; вони можуть всмоктуватися епітелієм слизової оболонки тонкої кишки в негідролізованій формі \ і розщеплюються до гліцерину та жирних кислот всередині ентероцитів.
Всмоктування продуктів перетравлення ліпідів
Складна суміш продуктів гідролізу ліпідів, зазначена вище, утворює ліпідні міцели, які можуть абсорбуватися слизовою оболонкою кишечника, і практично всі жири харчових продуктів надходять до грудної лімфатичної протоки завдяки всмоктуванню цих міцел. Проникнення ліпідних міцел всередину ентероцитів відбувається шляхом піноцитозу або дифузії окремих ліпідних молекул через апікальну мембрану клітин.
Порушення процесів перетравлення ліпідів
Порушення гідролізу та всмоктування харчових ліпідів у кишечнику супроводжуються розвитком стеатореї — наявності збільшеної кількості жирів у фекальних масах.
Розрізняють такі види порушень перетравлення ліпідів у кишечнику людини (А.Ш. Бышевский, О.А. Терсенов, 1994):
1) дефіцит панкреатичної ліпази, що спричинений захворюваннями підшлункової залози — панкреатична стеаторея;
2) дефіцит жовчі в кишечнику, пов’язаний з захворюваннями печінки або жовчних шляхів — гепатогенна стеаторея;
3) пригнічення ферментних систем ліполізу та ресинтезу триацилгліцеролів у кишечнику при його захворюваннях — ентерогенна стеаторея.