Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ I. БІОМОЛЕКУЛИ ТА КЛІТИННІ СТРУКТУРИ
ГЛАВА 3. НУКЛЕЇНОВІ КИСЛОТИ. НУКЛЕОТИДИ
3.3. БУДОВА, ВЛАСТИВОСТІ ТА БІОЛОГІЧНІ ФУНКЦІЇ ДНК
Біологічні функції ДНК:
1. Збереження спадкової інформації.
Кількість ДНК у соматичних та статевих клітинах організму людини є сталою величиною, яку ці клітини отримують у процесах запліднення батьківських гамет та подальшого поділу зиготи.
2. Передавання генетичної інформації нащадкам.
Подвоєння молекул ДНК у процесі реплікації та передавання нащадкам копій материнських молекул є основою консерватизму спадковості, збереження протягом багатьох поколінь основних біологічних ознак виду.
3. Реалізація генетичної інформації.
Ця біологічна функція здійснюється за рахунок передачі закодованої в ДНК інформації молекулам інформаційних (матричних) РНК (транскрипції) та подальшої розшифровки цієї інформації при синтезі білків (трансляції).
Сукупність зазначених біологічних функцій ДНК та механізмів їх реалізації' отримала назву — центральна догма молекулярної біології (Ф. Крік) (рис. 3.1):
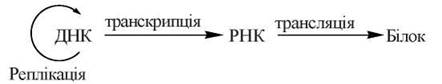
Рис. 3.1. Центральна догма молекулярної біології.
Експериментальне доведення генетичної ролі ДНК (феномен трансформації)
У 1928 р. англійським мікробіологом Ф. Гріффітом (F. Griffith) при вивченні двох штамів пневмококів Streptococcus pneumoniae — патогенного, що викликає пневмонію в людини та мишей (капсульної S-форми), та непатогенного мутанта (безкапсульної R-форми) — було відкрито явище трансформації. Вона полягала в можливості перетворення непатогенної R-форми в патогенну S-форму:
![]()
Гріффіт встановив, що трансформація пневмококів відбувається за умов взаємодії в організмі піддослідних тварин (мишей) вбитої нагріванням S-форми (патогенної) з живою непатогенною R-формою. Був зроблений висновок, що у вбитих нагріванням вірулентних клітинах пневмококів (штам S) присутній певний трансформуючий фактор, який, проникаючи в живі невірулентні клітини (штам R), змінює біологічні властивості останніх, надаючи їм властивість патогенності, до того ж ця властивість є спадковою:
![]()
Пізніше, в 1944 р., групою дослідників із Рокфелерівського інституту (США) — О. Евері, К. Мак-Леодом та М.Мак-Карті (O. Avery, C. McLeod, M. McCarthy) при дослідженні хімічної природи екстрактів патогенних пневмококів, які спричиняють трансформацію, було доведено, що трансформуючим фактором пневмококів є клітинна ДНК:
![]()
Таким чином, у результаті досліджень О. Евері та співавторів генетичне поняття «фактор спадковості», або «ген» уперше набуло конкретного молекулярного змісту — ним виявилась дезоксирибонуклеїнова кислота.
Молекулярна маса і розміри молекул ДНК
Молекулярна маса (м.м.) дезоксирибонуклеїнових кислот суттєво варіює в різних біологічних об’єктах: вірусах, прокаріотичних та еукаріотичних клітинах.
Точному визначенню молекулярної маси різних зразків ДНК перешкоджає гідродинамічна ламкість гігантських молекул нуклеїнових кислот, особливо у вищих організмів, які при спробі виділити їх в інтактному стані руйнуються на більш короткі фрагменти. Крім того, ДНК багатьох об’єктів має складну молекулярну організацію і становить широкий спектр різних полінуклеотидних конформацій: лінійні одноланцюгові та дволанцюгові молекули, кільцеві одноланцюгові та дволанцюгові молекули, суперспіралізовані структури.
Утім, застосування сучасних фізико-хімічних методів дослідження та електронної мікроскопії дозволило встановити, що молекулярна маса ДНК (при розрахунку на один полінуклеотидний ланцюг) складає в середньому діапазон від 106 до 1011 дальтон (Д).
ДНК вірусів і прокаріотів
Найменшу молекулярну масу та довжину молекули мають ДНК найпростіших живих утворень — вірусів, зокрема вірусів бактерій (бактеріофагів). Наприклад, м.м. одного з найменших за розміром бактеріофагів — фага —X 174 — становить (за даними різних досліджень) 1,6 · 106 (або 3,4 · 106) Д.
У прокаріотичних клітинах мікроорганізмів кількість ДНК та її молекулярна організація значно вищі, ніж у вірусів. Зокрема, ДНК кишкової палички E. Coli є ковалентно замкненим дволанцюговим кільцем з м.м. 1,9 · 109 (або, за іншими даними, 26 · 109) Д. Відповідно до зростання складності біологічної організації, при переході від вірусів до прокаріотів зростає і кількість нуклеотидних пар у дволанцюгових молекулах ДНК.
ДНК еукаріотичних клітин
У клітинах без’ядерних прокаріотів міститься одна молекула ДНК, яка розташована в спеціальній зоні цитоплазми — нуклеоїді. ДНК еукаріотів розміщена в ядрі й, поза фазами клітинного поділу, входить до складу аморфного нуклеопротеїнового утворення — ядерного хроматину. В період підготовки до мітозу (у фазі S клітинного циклу) відбувається подвоєння ДНК із подальшою конденсацією хроматину й утворенням цитологічних структур — хромосом, — в яких сконцентрований ядерний генетичний матеріал клітини.
За рахунок подвоєння (реплікації) молекул ядерної ДНК у фазі S у кожній соматичній клітині утворюється диплоїдний набір генетичного матеріалу, що забезпечує його рівномірний поділ між двома дочірніми клітинами. Таким чином, в період метафази клітинного циклу (період, що передує розходженню в дочірні клітини генетичного матеріалу) кожна хромосома еукаріотичної клітини містить дві повністю ідентичних молекули ДНК. Кількість хромосом в еукаріотичних клітинах специфічна для біологічного виду. Так, клітини улюбленого об’єкта класичної генетики — плодової мушки Drosophila melanogaster — мають по 8 хромосом, соматичні клітини людини — 46 хромосом. Молекулярна маса ДНК із хромосом людського організму дорівнює в середньому 1,6 · 1011 Д, що відповідає 2,4 · 109 парам азотистих основ. Фізична довжина розгорнутих молекул ДНК із клітин еукаріотів досягає декількох сантиметрів.
Молекулярні параметри розмірів ДНК деяких представників вірусів, прокаріотів та еукаріотів подано в табл. 3.3.
Таблиця 3.3. Молекулярна маса і розміри молекул ДНК із різних біологічних об’єктів
Біооб’єкт |
Молекулярна маса, Д |
Кількість нуклеотидних пар |
Довжина молекули |
Віруси |
|||
Вірус поліоми |
3 · 106 |
4,6 · 104 |
1,1 мкм |
Бактеріофаг λ |
3,3 · 107 |
5 · 104 |
13 мкм |
Бактеріофаг Т4 |
1,3 · 108 |
2 · 105 |
50 мкм |
Прокаріоти |
|||
Haemophilus influenzae |
8 · 108 |
1,2 · 106 |
300 мкм |
Escherichia coli |
1,9 · 109 |
3 · 106 |
1 мм |
Еукаріоти |
|||
Drosophila melanogaster |
4,3 · 1010 |
6,5 · 107 |
2 см |
Хромосоми людини |
1,6 · 1011 |
2,4 · 109 |
8,2 см |
Мітохондрії миші |
9,5 · 106 |
1,4 · 104 |
5 мкм |
Як свідчать дані таблиці 3.3, в цілому існує пряма пропорційність між зростанням еволюційного рівня, біологічної складності живих організмів та кількістю генетичного матеріалу, вираженого в чисельності нуклеотидних пар та, відповідно, молекулярній масі молекул ДНК. Значно менші молекулярні розміри та складність нуклеотидної організації ДНК мітохондрій тільки підтверджують існуючу теорію про походження цих органел із примітивних прокаріотів.
Вторинна структура
Вивчення нуклеотидного складу молекул ДНК із різних біологічних об’єктів показало, що, незалежно від джерела походження (бактеріальні, рослинні, тваринні організми), всі ДНК мають певні кількісні взаємовідносини між вмістом пуринових та піримідинових нуклеотидів. Згідно з цими закономірностями (правилами Чаргафа), у складі ДНК:
1) сума пуринових основ дорівнює сумі піримідинових основ, тобто:
А + Г = Т + Ц, або
2) кількість 6-аміногруп дорівнює кількості 6-кетогруп (за хімічною номенклатурою Фішера);
3) вміст аденіну дорівнює вмісту тиміну, а вміст гуаніну дорівнює вмісту цитозину (правило еквівалентності):
А = Т, Г = Ц.
Зазначені кількісні взаємовідношення між азотистими основами, а також результати вивчення будови молекул ДНК методом рентгеноструктурного аналізу (М. Уілкінс), дозволили американському біохіміку Джеймсу Уотсону та англійському фізику Френсису Кріку, що працювали в Кембриджському університеті, запропонувати просторову модель структури молекули ДНК у вигляді подвійної спіралі.
Згідно з моделлю Уотсона-Кріка, молекула ДНК складається з двох ланцюгів, що утворюють правообертаючу спіраль, в якій обидва полінуклеотидні ланцюги закручені навколо центральної осі; при цьому два полінуклеотидні ланцюги в молекулі ДНК антипаралельні (рис. 3.2, 3.3).
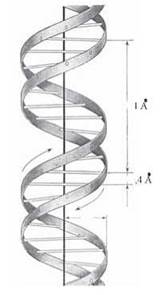
Рис. 3.2. Схематичне зображення двоспіральної молекули ДНК.
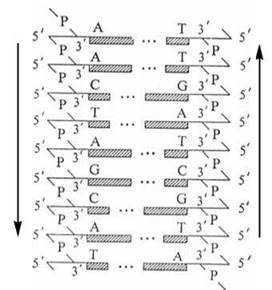
Рис. 3.3. Антипаралельність полінуклеотидних ланцюгів в молекулі ДНК.
Стабілізація подвійного ланцюга здійснюється за рахунок водневих зв’язків, що утворюються між протилежно розташованими, так званими комплементарними (додатковими), азотистими основами (аденіном і тиміном та гуаніном і цитозином, відповідно), що пояснює зазначені вище емпіричні правила Чаргафа.
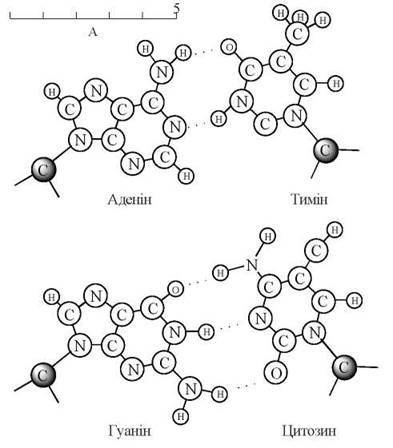
Крім водневих зв’язків, стабільність молекули ДНК підтримується також у результаті взаємодій між п-електронними хмарами гетероциклів азотистих основ, що розміщені один під одним вздовж осі спіралі — так звані «стекінг-взаємодії».
Структурні особливості подвійної спіралі: діаметр спіралі — 20 μ; відстань між азотистими основами впродовж осі спіралі — 3,4 Å; спіральна структура повторюється з інтервалом у 34 Å, тобто через 10 нуклеотидних пар.
Зазначені структурні особливості стосуються запропонованої Уотсоном і Кріком В-форми молекули ДНК (рис. 3.4). Разом із тим, залежно від взаємодії з різною кількістю молекул води та катіонами, ДНК набуває інших структурних форм: А, С та Z, які можуть відповідати певним фізіологічним умовам та взаємодії ДНК із білками ядерного хроматину (С-форма).
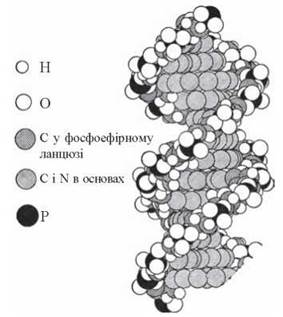
Рис. 3.4. В-форма молекули ДНК.
Третинна структура
У живій клітині подвійна спіраль, що становить вторинну структуру ДНК, не має вигляду розгорнутої молекули, а додатково згорнута в просторі, утворюючи певні третинні структури — суперспіралі.
У суперспіралізованому стані молекули ДНК у комплексі з певними клітинними білками входять до складу нуклеоїду прокаріотів та ядерного хроматину еукаріотів. Завдяки суперспіралізації довгі молекули ДНК формують компактні утворення, зокрема хромосоми ядра. Так, у результаті компактизації ядерна молекула ДНК клітин організму людини, що становить приблизно 8 см, вміщається в хромосомі довжиною 5 нм.
Фізико-хімічні властивості
Реакційноздатність
Усі полінуклеотиди, ДНК зокрема, є сильними багатоосновними кислотами з низьким значенням рК. Кислотність ДНК обумовлена вторинними фосфатними групами, що при рН > 4 повністю іонізовані.
Завдяки кислотним властивостям і наявності на своїй поверхні негативних зарядів молекули ДНК при фізіологічних значеннях рН активно реагують і утворюють комплекси з катіонами:
- поліамінами (спермідином, сперміном);
- основними білками (гістонами, протамінами);
- катіонами металів (Са2+, Mg2+, Fe2+).
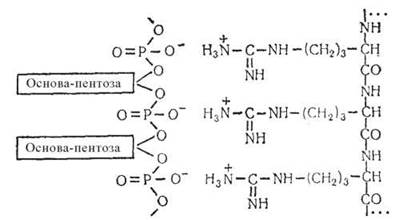
Рис. 3.5. Схема взаємодії поліїзуклеотидного ланцюга ДНК з основними білками.
В’язкість та оптична активність
Висока молекулярна маса і велика довжина молекул ДНК зумовлюють високу в’язкість навіть дуже розбавлених їх розчинів. В’язкість молекул ДНК у розчині залежить від їх конформації і суттєво змінюється за умов денатурації та ренатурації (див. нижче), що дозволяє використовувати віскозиметричні методи для дослідження кінетики цих процесів.
Завдяки впорядкованій вторинній структурі молекули ДНК є оптично активними, тобто вони здатні обертати площину поляризованого світла. Оптична активність розчинів ДНК також застосовується з метою реєстрації конформаційних змін молекул.
Поглинання в УФ-ділянці
Азотисті сполуки (та відповідні нуклеотиди), що входять до складу нуклеїнових кислот ДНК і РНК, мають властивість поглинати ультрафіолетове світло при 260 нм.
За умов утворення полінуклеотидів взаємний вплив паралельно розташованих по довжині молекули ДНК пар азотистих основ супроводжується певним зниженням УФ -поглинання. Таким чином, поглинання при 260 нм нативної' молекули ДНК дещо нижче (в середньому на 40 %) від відповідного поглинання суми азотистих основ, що входять до складу полінуклеотиду — гіпохромний ефект. При порушенні високовпорядкованої двоспіральної конформації ДНК та структурних взаємовідносин між азотистими основами спостерігається гіперхромний ефект, тобто зростання поглинання розчинів молекул ДНК при 260 нм, що дозволяє досліджувати процес денатурації.
Денатурація
Денатурація ДНК — це порушення нативної двоспіральної конформації молекул ДНК та їх упорядкованого просторового розташування з утворенням невпорядкованих одноланцюгових клубків. За умов денатурації ковалентні зв’язки в ДНК зберігаються, проте відбувається розкручування подвійної спіралі з втратою специфічних взаємодій між азотистими основами. Ренатурація — відновлення нативної вторинної конформації ДНК, що спостерігається за певних умов.
Денатурація ДНК супроводжується гіперхромним ефектом та зменшенням в’язкості її розчинів (рис. 3.6).
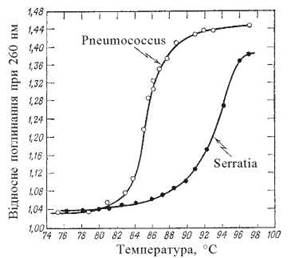
Рис. 3.6. Гіперхромний ефект у процесі денатурації ДНК мікроорганізмів.
Нуклеїнові кислоти, що піддані процесу денатурації, втрачають свої біологічні властивості.
Молекулярною основою денатурації молекул ДНК є руйнування водневих зв’язків між комплементарними азотистими основами А-Т та Г-Ц, відповідно.
Структурні зміни в молекулах ДНК, що призводять до їх денатурації, відбуваються внаслідок:
- різких змін рН у кислий або лужний бік;
- нагрівання розчинів ДНК до певних температур.
Термічна денатурація ДНК отримала назву процесу плавлення. Для кожного типу молекул ДНК (тобто молекул, що мають певний нуклеотидний склад) характерна відповідна температура денатурації, що позначається як температура (точка) плавлення — Тm (від англ. melting — плавлення). Тm різних зразків ДНК залежить від співвідношення в їх складі G-C- та A-T-пар.
Співвідношення між G-C- та A-T-парами є важливим показником нуклеотидного складу молекул ДНК з різних біологічних об’єктів. Оскільки між гуаніном та цитозином в складі двоспіральної молекули ДНК утворюється три водневих зв’язки (на відміну від двох — між аденіном та тиміном), термічна дисоціація пари G-C потребує більших затрат енергії, тобто відбувається при більш високих температурах, ніж руйнування пари A-T. Виходячи з цього, температура плавлення молекул ДНК прямо пропорційна вмісту в них G-C-пар, що дозволяє використати визначення Тm як показника нуклеотидного складу ДНК.