Підручник - БІОЛОГІЧНА ХІМІЯ - Губський Ю.І. - 2000
Розділ III. МЕТАБОЛІЗМ ОСНОВНИХ КЛАСІВ БІОМОЛЕКУЛ
ГЛАВА 16. МЕТАБОЛІЗМ ЛІПІДІВ. III. ОБМІН ХОЛЕСТЕРИНУ. ТРАНСПОРТ ЛІПІДІВ
16.2. БІОТРАНСФОРМАЦІЯ ХОЛЕСТЕРИНУ
Холестерин, що синтезувався в організмі, або надійшов із продуктами харчування, підлягає численним метаболічним перетворенням — біотрансформації, в результаті чого утворюються біологічно активні сполуки стероїдної природи, а також створюються умови для екскреції надлишків стеролу.
Першим етапом перетворення вільного холестерину є утворення його ефірів — холестеридів; збільшення активності реакцій біосинтезу холестерину супроводжується зростанням швидкості його етерифікації.
Етерифікація холестерину
Переважна частина холестерину тканин і близько 65 % холестерину плазми етерифіковано вищими жирними кислотами в положенні С-3. Синтез ефірів холестерину в плазмі крові та в клітинах відбувається за різними механізмами.
1. Зовнішньоклітинна етерифікація холестерину здійснюється ферментом лецитин (фосфатидилхолін)-холестерин-ацилтрансферазою (ЛХАТ) плазми. ЛХАТ каталізує реакцію перенесення ацильного залишку з 2-го (β-) положення фосфатидилхоліну на гідроксильну групу холестерину:
![]()
Для етерифікації холестерину використовуються ненасичені жирні кислоти: лінолева С182 (переважно) та олеїнова С181. Найбільш активно реакція відбувається в ліпопротеїнах крові ЛПВЩ (див. розділ 16.3).
2. Внутрішньоклітинна етерифікація холестерину перебігає за участю ацил-КоА-холестерин-ацилтрансферази (АХАТ):
![]()
АХАТ використовує для етерифікації холестерину ацил-КоА різної природи, найбільш активно — залишки олеїнової кислоти. Фермент локалізований у мембранах ендоплазматичного ретикулума (мікросомній фракції) клітин печінки та інших органів — надниркових залоз, кишечника, шкіри тощо. Фізіологічною функцією АХАТ є створення внутрішньоклітинних резервів холестерину, який після деетирифікації спеціальними естеразами, може використовуватися для синтезу жовчних кислот, стероїдних гормонів наднирникових залоз, секс-гормонів, вітаміну D3 та його похідних.
Шляхи біотрансформації холестерину
Біотрансформація холестерину в інші біологічно активні сполуки стероїдної природи здійснюється за рахунок введення в молекулу стеролу додаткових гідроксильних груп та реакцій модифікації в бічному ланцюзі. Реакції окисного гідроксилювання стероїдів каталізуються ферментами монооксигеназами (оксигеназами мішаної функції).
Сумарне рівняння процесу окисного гідроксилювання холестерину (RH), який відбувається при синтезі його біологічно активних похідних, має вигляд:
![]()
Процес перебігає за участю цитохрому Р-450 у мембранах ендоплазматичного ретикулума гепатоцитів («мікросомальне окислення») або в мітохондріях наднирникових залоз та клітин статевих залоз.
1. Біосинтез жовчних кислот.
У гепатоцитах холестерин перетворюється на жовчні кислоти — важливі компоненти жовчі, що беруть участь у перетравленні харчових жирів у кишечнику людини і тварин.
Жовчні кислоти є гідроксильованими похідними холанової кислоти; до них належать такі сполуки: холева (3,7,12-триоксихоланова), дезоксихолева (3,12-діоксихоланова), хенодезоксихолева(3,7-діоксихоланова) та літохолева (3-оксихоланова) кислоти.
Холева та хенодезоксихолева кислоти (первинні жовчні кислоти) утворюються в печінці при гідроксилюванні циклопентанпергідрофенантренового циклу та частковому окисленні в боковому ланцюзі молекули холестерину.
Першим етапом у біосинтезі жовчних кислот є 7α-гідроксилювання холестерину, що каталізується ферментом мембран ендоплазматичного ретикулума 7α-гідроксилазою, який є однією з ізоформ цитохрому Р-450 та функціонує за участю НАДФН, кисню та вітаміну С (аскорбінової кислоти).
7α-Гідроксилаза — регуляторний фермент, що є активним у фосфорильованій і малоактивним — в дефосфорильваній формі. Жовчні кислоти — кінцеві продукти цього метаболічного шляху — пригнічують активність ферменту за принципом негативного зворотного зв’язку.
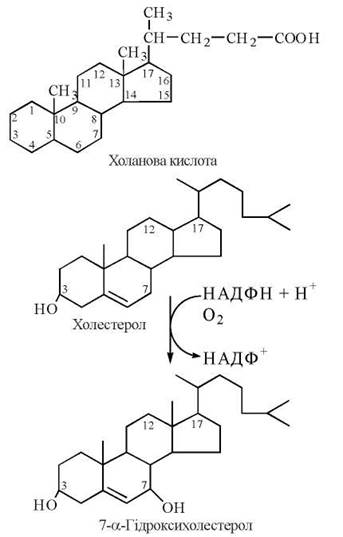
Після утворення 7α-гідроксихолестеролу шлях біосинтезу жовчних кислот дихотомічно розгалужується: одна з гілок веде до утворення холевої кислоти, друга — хенодезоксихолевої. Ці сполуки надходять із гепатоцитів у жовчні капіляри і депонуюються в жовчному міхурі, надходячи з нього до порожнини дванадцятипалої кишки. При дії ферментів мікроорганізмів, що містяться в кишечнику, утворюються вторинні жовчні кислоти — дезоксихолева та літохолева (рис. 16.2):
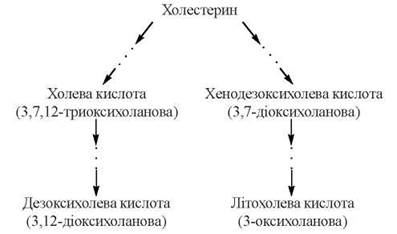
Рис. 16.2. Схема утворення жовчних кислот із холестерину.
Основним, щодо кількості, представником жовчних кислот у жовчі людини є холева кислота, яка бере участь в емульгуванні жирів у кишечнику у вигляді натрієвої та калієвої солей її кон’югованих форм — глікохолевої та таурохолевої кислот. Глікохолат і таурохолат містять в своїй структурі гідрофільні (радикали гліцину та таурину) та гідрофобні (стероїдне ядро) молекулярні групи, і завдяки своїй амфіпатичній будові є високоактивними детергентами, що необхідні для емульгування жирів у кишечнику.
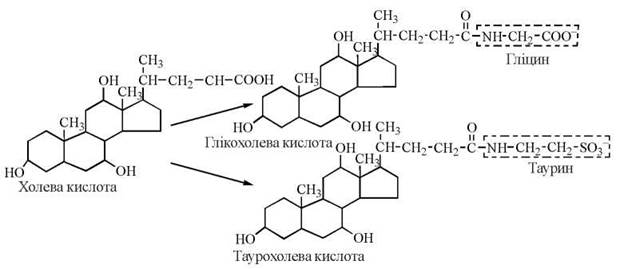
Холева кислота та її кон’юговані форми — глікохолат і таурохолат.
2. Біосинтез стероїдних гормонів
Стероїдні гормони містять у своєму складі 21 (кортикоїди, прогестерон) і менше (19 — андрогени, 18 — естрогени) атомів вуглецю, тому їх утворення з С27-стероїду холестерину включає, крім окисного гідроксилювання, і розщеплення вуглеводневого бічного ланцюга, реакції окислення, відновлення та ізомеризації.
Першим етапом на шляху синтезу з холестерину стероїдних гормонів надниркових залоз (кортикостероїдів) є утворення С21-стероїду прегненолону — безпосереднього попередника прогестагену прогестерону (С21), який у клітинах надниркових залоз перетворюється на кортикостероїди (С21): глюкокортикоїд кортизол та мінералокортикоїд альдостерон.
Гормони чоловічих та жіночих статевих залоз також утворюються з холестерину через стадію прегненолону та прогестерону, який у цих органах перетворюється в 17-α-гідроксипрогестерон — попередник андрогену (С19) — тестостерону та естрогенів (С18) — естрону та естрадіолу.
3. Біосинтез вітаміну D3.
Перетворення холестерину у вітамін D3 — холекальциферол — потребує розщеплення кільця циклопентанпергідрофенантрену з утворенням провітаміну D3, який підлягає реакціям окисного гідроксилювання з утворенням біологічно активної форми вітаміну — 1,25-дигідроксихолекальциферолу (кальцитріолу). Більш детально процес біосинтезу вітаміну D3 та його активних форм буде розглянутий у главі 25.
Схему біотрансформації холестерину з утворенням жовчних кислот та біологічно активних стероїдів подано на рис .16.3.
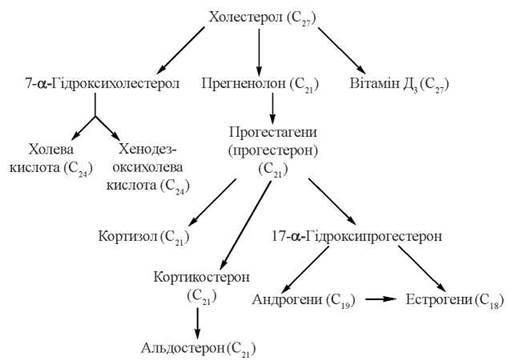
Рис. 16.3. Метаболічна карта біотрансформації холестерину.
Екскреція холестерину з організму
За рахунок ендогенного синтезу та всмоктування з кишечника пул холестерину в організмі міг би щодобово збільшуватися приблизно на 1 г. Цьому протидіють реакції виведення холестерину з обміну шляхом його перетворення в інші стероїди та екскреції з організму з фекаліями.
Шляхи екскреції холестерину:
1. Виведення у формі жовчних кислот. В організмі постійно циркулює (в процесі ентерогепатичної циркуляції) близько 3-5 г жовчних кислот; певна частина цієї кількості (до 500 мг/добу) не всмоктується і виводиться з організму.
2. Виведення у формі копростанолу — продукту, що утворюється з холестерину в нижньому відділі кишечника під дією ферментів кишкової мікрофлори. Цим шляхом екскретується та частина холестерину, яка не абсорбується в кишечнику.
3. Виведення у формі нейтральних стероїдів — кінцевих метаболітів стероїдних гормонів.
Порушення динамічної рівноваги між процесами, що постачають холестерин та виводять його з організму, призводить до надмірного накопичення стеролу в клітинах та екстрацелюлярних просторах — клініко-біохімічного синдрому холестеринозу (Ю.М. Лопухин, 1983), проявами якого є сімейні (спадкові) гіперхолестеринемії та атеросклероз.