ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 4. ЕКОЛОГО-БІОХІМІЧНІ ВЗАЄМОДІЇ РОСЛИН І ТВАРИН
4.3. Гормональні взаємодії між рослинами і тваринами
У тварин гормони зазвичай виробляються ендокринними залозами, а потім транспортуються до місця дії системою кровообігу. Більшість гормонів тварин є пептидами або стероїдами. Водночас у рослинах синтезувати гормони має здатність багато клітин, місце синтезу змінюється з розвитком рослини, гормони переносяться на відносно малу відстань. Гормони рослин мають різну хімічну структуру — містять пуринові основи (цитохініни) або амінокислоти (ауксини), можуть бути терпеноїдами (дорміни, гібереліни) тощо. Разом з тим, установлено, що рослини здатні продукувати ряд гормонів тварин.
Статеві гормони тварин, які продукуються рослинами. До гормонів тварин, які виділяються рослинами, належать жіночі та чоловічі гормони ссавців, у тому числі людини. Одна з гіпотез стосовно їхніх функцій — вони здатні запобігати поїданню рослин ссавцями. У тварин гормональна система ретельно збалансована. Тому екзогенне надходження статевих гормонів може серйозно впливати на розмноження. Так, уживання в їжу цибулин тюльпанів зумовлює порушення менструального циклу та недостатню овуляцію в жінок. Іншими джерелами гормонів, які впливають на еструс у жінок, є часник, овес, ячмінь, райграс, кава, соняшник, петрушка, бульби картоплі.
За іншою гіпотезою статеві гормони ссавців у рослин іноді можуть утворюватися випадково як продукти метаболізму, що приводять до синтезу важливіших стеролів рослин, або ж їхня роль пов’язана з процесами розвитку, цвітіння та проявів статі в рослин. Так, якщо статеві гормони ссавців, у тому числі людини, вносити екзогенно в середовище проростання рослин або в самі рослини, то: естрогени стимулюють проростання насіння та ріст рослин, а також сприяють розвитку квітів; тестостерон збільшує кількість жіночих зародків.
Статеві гормони людини, які продукуються рослинами, наведено в табл. 4.5, а їхню структуру — на рис. 4.10.
Таблиця 4,5
СТАТЕВІ ГОРМОНИ ЛЮДИНИ, ВИЯВЛЕНІ В РОСЛИНАХ
(за Харборном, 1985, зі змін.)
Гормон |
Рослина-продукт |
Естрон |
Фінікова пальма Phoenix dactylifera (насіння, пилок); Гранатове дерево Punica granatum (насіння) |
Естріол |
Верба Salix (квіти); Яблуня Malus domestica (насіння) |
β-Естрадіол |
Квасоля Phaseolus vulgaris (насіння) |
Тестостерон |
Сосна Pinus sylvestris (пилок) |
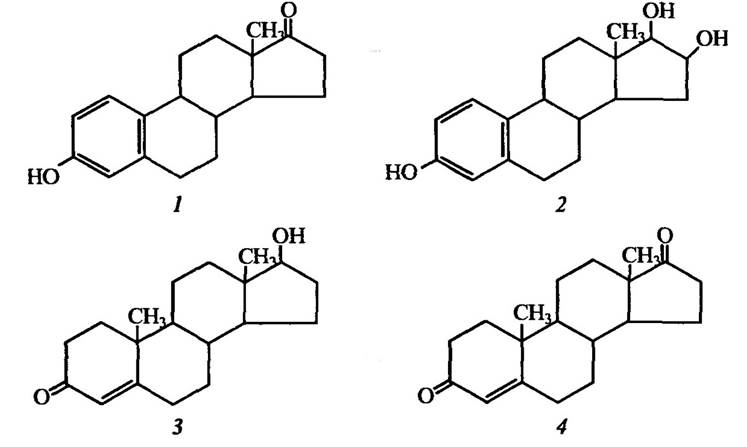
Рис. 4.10. Структура статевих гормонів людини, які виявлені в рослинах:
естрогени — естрон (1) і βр-естрадіол (2); андрогени — тестостерон (5) і андростендіон (4)
Крім насіння та пилку, статеві гормони людини виявлено також, хоча і в дуже малій кількості, у листі та корінні, зокрема дерева Punica granatum.
У деяких випадках, а можливо, і в усіх рослинах, містяться не самі гормони тварин, а сполуки, які імітують їхню дію. Так, із коренів рослини родини бобових Pueraria mirifica виділено мірестрол (рис. 4.11), структура якого близька до жіночого гормону естрону. Ця сполука за перорального вживання викликає аборт у вагітних жінок.
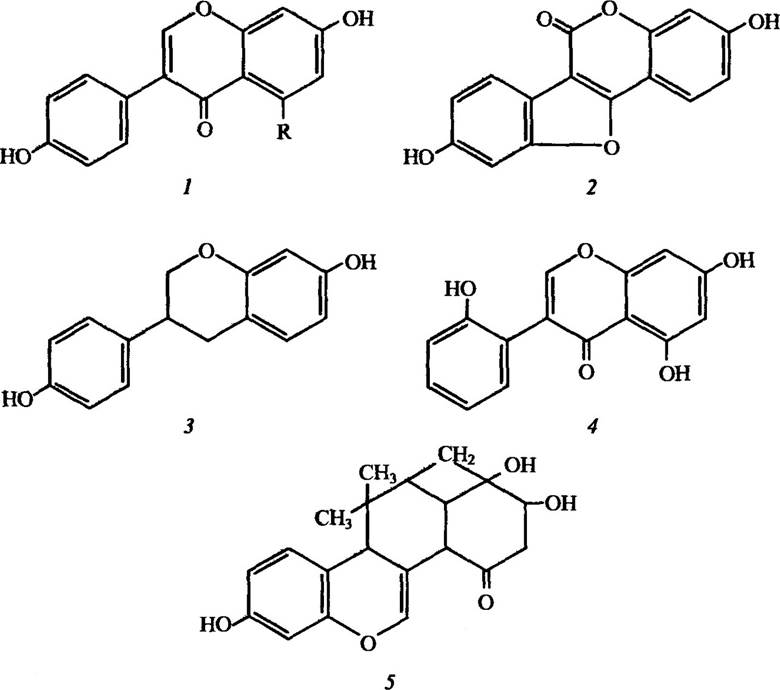
Рис. 4.11. Структура деяких імітаторів естрогенів:
1 — геністеїн (R=OH), даідзеїн (R=H), формононетин (R=H, СН3 при 4-ОН); 2 — куместрол;
3 — еквол; 4 — ізогеністеїн; 5 — мірестрол
Установлено, що деякі ізофлавоноїди здатні здійснювати екстрогенну дію на ссавців. Так, із конюшини підземної Trifolium subterraneum було виділено геністеїн і формононетин, які спричинюють стерильність овець.
З люцерни Medicago sativa та конюшини повзучої Trifolium repens виділено куместрол, активність якого майже в 30 разів більша, ніж геністеїну. Ця сполука здатна в результаті деметилювання та відновлення в організмі тварин перетворюватися в еквол.
Ізофлавони впливають не тільки на ссавців, а й на шахів. Так, самки перепелів відкладають менше яєць у кладці, якщо вони харчуються на пасовищах, багатих бобовими з високим рівнем ізофлавонів у листі.
На плодючість деяких тварин можуть впливати навіть простіші, ніж ізофлавони сполуки. Репродуктивну функцію гризунів Microtus montanus пригнічують фурулова та n-кумарова кислоти, а також їхні вінільні аналоги. Ці сполуки здатні викликати зменшення маси матки, затримування розвитку фолікул і, зрештою,—зниження плодючості.
Ювенільні гормони та линяння в комах. Найпереконливіші докази гормональної взаємодії між рослинами та тваринами отримано при дослідженні відкритих в рослинах гормонів линяння та ювенільних (пат. juvenalis — статевонезрілий) гормонів.
У метаморфозі комах гормони необхідні для регуляції онтогенезу, який охоплює кілька стадій за участю гормонів линяння (ГЛ) та ювенільного гормону (ЮГ)— від личинки до дорослої особини (імаго). Вони необхідні для ініціації перетворень у процесі розвитку комах, які спрощено можна представити так:

де n — кількість личинкових стадій (5 і більше).
Ювенільні гормони (рис. 4.12), зокрема ювабіон, його дегідрована похідна дегідроювабіон, ювоцимени та ін. необхідні тільки для переходу личинки з першої стадії в другу. Вони синтезуються парою залоз, які розміщені біля голови комах, і потрапляють у кров. За структурою вони сесквитерпени. Прекоцен 1 і прекоцен 2 викликають передчасний розвиток, личинки за їх наявності не проходять одну або кілька стадій і перетворюються в неповноцінну дорослу форму. Ці сполуки інгібують синтез ювенільних гормонів тільки після того, як у залозах комах вони перетворюються в епоксидну форму.
Сполуки рослин з активністю ювенільних гормонів синтезуються як засіб самозахисту від комах, оскільки вони здатні затримувати розвиток комах, а іноді — спричиняти їх загибель. Можливість здійснювати контроль за ростом комах за допомогою подібних речовин використовують під час штучного синтезу інсектицидів. Усю групу речовин, які пригнічують синтез ювенільних гормонів (блокують функціонування залоз), називають анатоксинами (див. розд. 2).
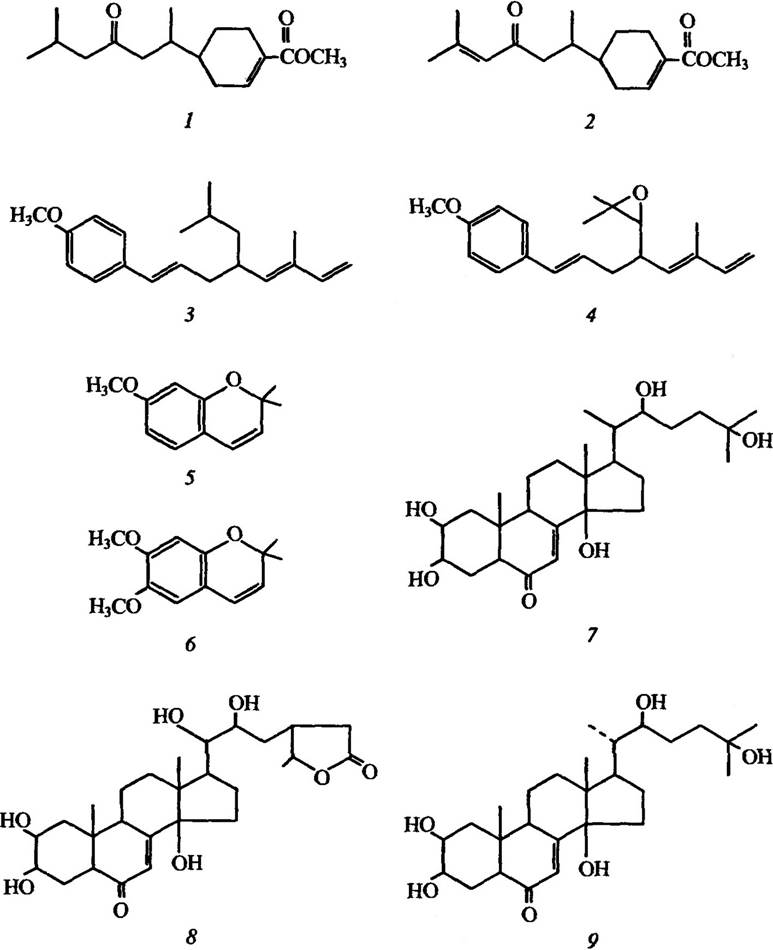
Рис. 4.12. Структура гормонів комах:
1 — ювабіон; 2 — дегідроювабіон; 3 — ювоцимен 1; 4 — ювоцимен 2; 5 — прекоцен 1; 6 — прекоцен 2;
7 — α-екдизон; 8 — цикастерол; 9 — β-екдизон (екдистерон)
Гормони линяння контролюють линяння зовнішньої оболонки кутикули на кожній стадії метаморфозу та необхідні на всіх етапах розвитку комах. Ці гормони продукуються в проторакальних залозах, у передньому грудному відділі комахи. Гормони линяння називають екдизонами (від лат. ekdu — знімати із себе, линяти і
вихід).
У складі гормонів линяння виявлено як α-екдизон, так і β-екдизон (рис. 4.12), який може утворюватися з α-екдизону. У різних членистоногих і ракоподібних були виявлені також подібні за структурою гормони линяння.
У рослинах виявлено близько 50 речовин, що проявляють активність гормонів линяння і які через їхню високу активність (для деяких майже у 20 разів порівняно з α- і β-екдизонами) порушують нормальний метаморфоз комах.
Екдизони (β-екдизон), які вперше було виявлено в листі тиса ягідного Taxus baccata, трапляються в папороті, голонасінних і значно рідше — у покритонасінних. У всіх рослин, які містять гормони линяння комах, виявлено Р-екдизон, інші гормони зустрічаються рідко. Разом з тим у рослинах помічено широкий набір речовин, які подібні за структурою до екдизонів і проявляють активність гормонів линьки. Представником таких речовин є цикастерол (рис. 4.12) з Ajuga remota, який порушує линяння в личинок совки Spodoptera frugiperda і коробкової молі Pectinophora gossypiella. Цей фітогормон викликає появу тришарової кутикули: нова наростала, а стара не скидалася. Наслідком цього стало порушення будови ротового апарату, у результаті личинки гинули від голоду. Плюмбіган з Plumbago capensis і азадирахтин з Melia azedarach та ще багато інших фітогормонів є не тільки інгібіторами линяння в комах, а й антифідантами.
Біологічна дія фітогормонів залежить від того, потрапили вони на верхні покриви тіла личинки чи в організм перорально. Показано можливість фітогормонів, які надійшли через рот з їжею, зазнавати подальшого гідроксилювання або гідрогенізації, відщеплення бокових ланцюгів, утворень кон’югатів тощо, що викликає їх детоксикацію.
Якщо характеризувати фітогормони линяння комах у цілому, то треба зупинитись ось на чому. Вони мають значно більшу активність і термін інактивації, ніж екдизони. Потрапивши в організм, вони спричиняють істотні порушення росту, стерильність і навіть смерть. Ці речовини не обов’язково мають зумовити загибель личинок комах. Навіть незначний вплив на метаморфозм і розмноження комах у більшості випадків є достатнім, щоб зробити рослину менш придатною для шкідників і контролювати в певних межах їхню чисельність на рослині. Деякі комахи (їхні личинки) здатні до детоксикації фітогормонів линяння, що потрапляють з їжею, але інші не можуть інтоксикувати їх своєчасно. Личинки комах меншою мірою здатні протидіяти таким речовинам у разі надходження їх в організм через кутикулу. Фітогормони линяння личинок комах потрапляють, головним чином, у двох груп відносно примітивних рослин — папороті та голонасінних, які рідше, як покритонасінні, зазнають нападу комах-шкідників. Отже, наявність фітогормонів линяння комах — захисний механізм, який був вироблений рослинами на певному етапі еволюції до появи або тільки на початку розвитку покритонасінних. У цих рослинах подібну функцію виконують алкалоїди та елагові таніни.