ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 8. СТІЙКІСТЬ ТА АДАПТАЦІЯ РОСЛИН І ТВАРИН
8.3. Стійкість та адаптація тварин до умов існування
8.3.2. Осморегуляція
Для оптимального функціонування організмів потрібен цілком визначений і відносно сталий склад рідин тіла. Живі організми здатні підтримувати необхідні для життєдіяльності концентрації розчинених речовин. Найважливішою за значенням для організму є вода. Згідно з домінуючим уявленням, саме у воді прибережних лагун зародилося життя. Роль, яку відіграє вода в біологічних системах, визначається її здатністю розчиняти речовини, стабілізувати біологічно важливі молекули, слабкою іонізацією, великим значенням захованої теплоти й пароутворенням, поверхневим натягом, бути субстратом у синтезі та гідролізі ряду речовин тощо. Вона визначає фізичні властивості клітин — їхній об’єм, внутрішньоклітинний тиск (тургор). Вода є основним середовищем, де відбуваються всі процеси життєдіяльності. Її вміст у більшості живих організмів становить 65—75 % загальної маси, а в деяких— до 98 % (медузи).
Вода формує оболонку навколо деяких сполук (наприклад, білків), що перешкоджає їхній взаємодії. Таку воду називають зв’язаною (структурованою), її частка становить 4—5 % загальної кількості води в організмі. Інша частина води (94—95 %) — це так звана вільна вода.
Необхідний вміст води в організмі підтримується надходженням її ззовні (для людини приблизно 2 л за добу) і невеликою кількістю води (0,3 л за добу), яка утворюється в процесі катаболізму речовин.
Вода складається з двох атомів гідрогену, які одинарним ковалентним зв’язком сполучаються з атомом оксигену. Електрони в молекулі води розподілені несиметрично, і кут між зв’язками Н-О-Н дорівнює 104,5°. Унаслідок цього вода має полярні властивості, а сама молекула становить собою диполь. Атом оксигену в молекулі води притягує електрони атома гідрогену. Унаслідок цього атом гідрогену стає електропозитивним, а оксигену — електронегативним. Між атомом оксигену й гідрогену сусідніх молекул виникають гід- рогенні зв’язки, які сприяють утворенню агрегатів (асоціатів) — кожна молекула вода може зв’язувати чотири сусідні, у результаті чого виникають пентанольні структури.
За здатністю розчинятися у воді органічні та неорганічні сполуки поділяють на розчинні у воді — гідрофільні (гр. hydro — вода і phileo — дружба), або полярні, та нерозчинні в ній — гідрофобні (гр. phobos — страх), або неполярні.
До гідрофільних сполук належить більшість кристалічних солей, наприклад натрію хлориду. Протилежні електричні заряди атомів гідрогену й оксигену притягують іони Na+ і Сl-, витягують їх із кристалічної ґратки. Вода розчиняє також багато сполук, що здатні іонізуватися. До таких речовин належать органічні сполуки, що містять, наприклад, карбонільні групи (альдегіди, кетони), гідроксильні (спирти) тощо.
Більшість вуглеводних сполук нерозчинні у воді. Це зумовлено тим, що між водою і цими сполуками виникають гідрофобні взаємодії— неполярні молекули оточуються найближчими молекулами води й формуються структури з обволіканням водою. Гідрофобна взаємодія має істотне значення для структурування біомембран, стабілізації білків та нуклеїнових кислот.
Значна кількість речовин є амфіпатичними, які містять як полярні, так і неполярні групи. У водневому середовищі наявність подібних груп спричиняє гідрофобний ефект — неполярні частки молекули намагаються об’єднатися, а молекули води відштовхуються. Це призводить до утворення міцел.
Підтримка належних концентрацій розчинених речовин в рідинах тіла тварин є основою підтримки гомеостазу (гр. homios — подібний і stasis — стан) — сукупності пристосувальних реакцій організму, спрямованих на подолання чи максимальне обмеження дії різних факторів довкілля або внутрішнього середовища, що здатні порушувати відносну сталість внутрішнього середовища організму. Залежно від навколишнього середовища концентрація води й розчинених у ній сполук здатні змінюватися і принциповим є місце мешкання організмів — морська або прісна вода, суходіл.
Водне середовище. Водою покрито 71 % поверхні Землі. В основному це вода океанів. Прісна вода становить приблизно 0,01 % об’єму морської води. Будь-яка вода містить розчинні в ній солі, гази, невелику кількість органічних сполук, різні домішки. У табл. 8.6 наведено іонний склад морської, а також річкової, жорсткої річкової та солоної води суходолу.
Таблиця 8.6
ІОННИЙ СКЛАД МОРСЬКОЇ І ПРІСНОЇ ВОДИ (г/кг води)
ІОН |
Морська вода |
Вода суходолу |
||
річкова |
жорстка |
солона |
||
Натрій |
10,933 |
0,39 |
6,13 |
640 |
Магній |
1,317 |
0,21 |
0,66 |
6 |
Кальцій |
0,414 |
0,52 |
5,01 |
32 |
Калій |
0,394 |
0.04 |
0,1 1 |
16 |
Хлор |
19,658 |
0,23 |
13,44 |
630 |
Сульфат |
2,744 |
0,21 |
1,40 |
54 |
Бікарбонат |
0,145 |
1,11 |
1,39 |
3 |
Морська вода містить 3,5 % солей. Іони в основному представлені натрієм і хлором, у значній кількості наявні магнам, сульфат і кальцій. Концентрація солей залежить від географічного положення,
припливу прісної води у прибережні райони тощо. Однак відносна кількість розчинних іонів у Світовому океані залишається незмінною.
У прісній воді, на відміну від морської, концентрація розчинних солей може бути різною. У дощову воду солі потрапляють із морської — її бризки випаровуються і частинки солі потоками повітря переносяться на суходіл із дощовою водою потрапляють на землю. Якщо вода тече по твердій нерозчинній породі, то в неї майже не потрапляють розчинені речовини й вона називається . Коли вода просочується в пористий вапняк, то в ній розчиняється відносно багато солей кальцію, і таку воду називають жорсткою.
У деяких місцях вода внутрішніх водойм містить багато солей — так звана солона вода. Солонуваті води трапляються в прибережних районах, де морська вода перемішується з прісною.
Солестійкість галофільних бактерій. У бактерій цитозоль і зовнішнє середовище їхнього мешкання мають однакову концентрацію солей у великому діапазоні солоності. Бактерії, які пристосувалися до великих концентрацій солей, не просто здатні функціонувати за цих умов, а й потребують цих кількостей солей для життєдіяльності. Одними з найсолестійкіших організмів є галофільні (гр. halos — сіль та philos — любити) бактерії. У табл. 8.7 наведено концентрації Na+ і K+ у негалофільних і галофільних бактеріях.
Таблиця 8.7
ВНУТРІШНЬОКЛІТИННА КОНЦЕНТРАЦІЯ СОЛЕЙ У РІЗНИХ БАКТЕРІЙ
(за Хочачкою, Семеро, 1977, зі змін.)
Бактерії |
Концентрація в культуральній рідині, моль · л-1 |
Внутрішньоклітинна концентрація, міліеквівалент/1 кг клітинної води |
||
NaCI |
КСl |
Na+ |
K+ |
|
Негалофіли: Staphylococcus aureus |
0,15 |
0,025 |
98 |
680 |
Salmonella typhi |
0,15 |
0,025 |
131 |
239 |
Помірні галофіли: Micrococcus halogenitrificans |
1,0 |
0,004 |
311 |
474 |
Vibrio costicolus |
1,0 |
0,004 |
684 |
221 |
Справді галофіли: |
||||
Sarcina morrhuae |
4,0 |
0,032 |
3170 |
2030 |
Halobacterium salinarium |
4,0 |
0,032 |
1370 |
4570 |
Зауважимо, що в галофільних бактерій існують у фізіологічному аспекті ті самі метаболічні шляхи, що й у негалофітів: синтез АТФ, реплікація генів і синтез білка тощо. Однак у них біологічні реакції відбуваються з урахуванням тих умов довкілля, у яких живуть ці екстремальні організми. Так, наприклад, НАДФ+-ізоцитратдегідрогеназа каталізує реакцію:
![]()
Ця ферментна система галофільних бактерій проявляє свою максимальну активність за концентрації Na+ або K+ 1,0 моль/л.
Аспартат-карбамоїлтрансфераза каталізує реакцію, яка є першим етапом біосинтезу піримідинів:
![]()
Для прояву активності цього ферменту в галофілів необхідна концентрація солей 3,0 — 5,0.
У галофільних організмів від оптимальної високої концентрації солей залежить функціонування клітинних органел. Так, наприклад, у рибосом 30 S і 150 S субодиниці стабільні за концентрації NaCl або КСl 1,0 моль/л, а утворення 70 S-субодиниці відбувається за 3,0 — 4,0 КСl. Амінокислоти рибосом галофільних мікроорганізмів відрізняються від амінокислотного складу рибосом інших бактерій і еукаріотичних клітин надзвичайно високим вмістом кислих амінокислот.
Установлено, що білки клітинної стінки й плазматичної мембрани галофільних бактерій мають кислий характер. Для їх стабілізації, як і для рибосом, також необхідні високі концентрації солей. Узагалі показано, що більшість білків (до 89 %) галофільних бактерій є кислими. Тому для нейтралізації негативних зарядів, які містяться на їхній поверхні, необхідні високі концентрації катіонів.
Основні способи осморегуляції. У багатьох морських і евригалинних(гр. eurys — широкий та gals — сіль) безхребетних, які здатні переносити великі коливання концентрації солей у воді, кров гіперосмотична, ступінь якої неоднаковий у різних тварин. У них зменшення солоності середовища мешкання супроводжується зниженням концентрації осмотично активних одиниць (неорганічних іонів, амінокислот та ін.) у крові та навпаки — збільшення концентрації солей у довкіллі викликає підвищення їхньої кількості у крові.
З’ясовано, що один із механізмів осмотичної адаптації — зміна концентрації внутрішньоклітинних амінокислот. Так, у ракоподібних (Cypridina) такими основними амінокислотами є гліцин, аланін, пролін і глутамінова кислота, хоча відносний внесок кожної з них у різних видів неоднаковий. У морських зірок (тип Echinodermata) в осморегуляції беруть участь гліцин і таурин, у кільчастого черва Arenicola — гліцин, аланін і глутамінова кислота. Наслідком адаптивних змін вмісту цих амінокислот є сталість об’єму клітин, і визначається кількістю осмотично активних речовин.
На початкових етапах регуляції об’єму клітин істотна роль належить глутаматдегідрогеназі, яка каталізує реакцію:
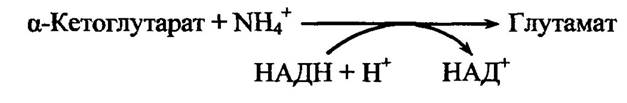
Глутаматдегідрогеназа активується неорганічними іонами. Ця реакція є головним шляхом утворення а-аміногруп з NH4+. Амінування а-кетокислот з використанням глутамату як донора а-аміногруп приводить до утворення інших амінокислот, у тому числі гліцину, аланіну та проліну.
Істотне значення в осморегуляції належить активному (із затратою енергії, зокрема АТФ) транспорту іонів мембранозв’язаними транспортними «насосами» — Na+, К+-АТФазі, Са2+-АТФазі, Н+- АТФазі та ін. Вторинний активний транспорт відбувається за рахунок мембранного електрохімічного потенціалу.
У регуляції внутрішньоклітинної концентрації Na+ суттєве місце належить локалізованій у плазматичній мембрані Na+, К+-АТФазі, яка переносить 3Na+ назовні і 2K+ всередину клітин. Активність цього ферменту залежить від фосфоліпідів мембран, які здатні перетворювати фермент із неактивної в активну форму. У сольовій залозі птахів, через яку в них виділяється надлишок Na+, активність Na+, K+- АТФази збільшується з підвищенням вмісту сульфатиду, до складу якого входять сфінгозин, 2-окситетракозанова кислота і галактоза, яка етерифікована сірчаною кислотою (рис. 8.1).
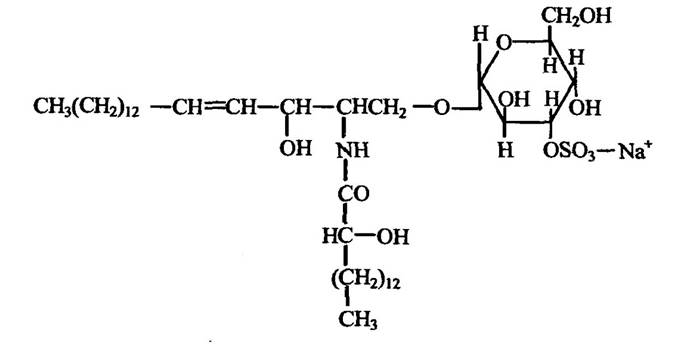
Рис. 8.1. Структура сульфатиду із сольової залози качки
Активність Na+, K+-АТФази найбільша в тих тканинах, головна функція яких полягає в перенесенні електролітів. Вона міститься і в інших тканинах тіла, але в меншій кількості. У кишечнику цей фермент переносить Na+ через кишкову стінку, у нирках він бере участь в абсорбації Na+, у нервовій тканині — у реполяризації мембрани після проведення електричного імпульсу тощо.
У регуляції концентрації Na+ можуть брати участь, крім К+, також інші іони, зокрема NH4+, Н+.
ОСОБЛИВОСТІ АДАПТАЦІЇ ТВАРИН ДО СОЛОНОСТІ ВОДИ. Для підтримування відносної іонної сталості рідин тіла тварин повинні регулювати як надходження в організм солей, так і їх виведення. Такі організми, яких називають осморегуляторами, мають притаманні їм системи осморегуляцїї. Існують також організми, у яких склад рідин відповідає складу навколишнього водного середовища, — осмоконформери.
Водні безхребетні тварини. Як правило, морські безхребетні тварини є осмоконформерами, але вміст деяких речовин, розчинних у рідинах тіла, відмінний від їхньої кількості у воді. Концентрацію найважливіших іонів у крові деяких безхребетних наведено в табл. 8.8.
Таблиця 8.8
КОНЦЕНТРАЦІЯ ІОНІВ У МОРСЬКІЙ ВОДІ ТА РІДИНАХ ТІЛА ДЕЯКИХ МОРСЬКИХ ТВАРИН
(за Шмідтом-Ніельсоном, 1982)
Вода або організм |
Іони, ммоль/кг води |
|||||
Na+ |
Mg2+ |
Са2+ |
K+ |
Сl- |
SO42- |
|
Морська вода |
478 |
54,3 |
10,5 |
10,1 |
558,4 |
28,8 |
Медуза (Aurelia) |
474 |
53,0 |
10,0 |
10,7 |
580 |
15,8 |
Поліхета (Aphrodite) |
476 |
54,0 |
10,5 |
10,5 |
557 |
26,5 |
Морський їжак (Echinus) |
474 |
53,5 |
10,6 |
10,1 |
557 |
28,7 |
Мідія (Mytilus) |
474 |
52,6 |
П,9 |
12,0 |
553 |
28,9 |
Кальмар (Loligo) |
456 |
55,4 |
10,6 |
22,2 |
578 |
8,1 |
Рівноногий рак (Ligia) |
566 |
20,2 |
34,9 |
13,3 |
629 |
4,0 |
Краб майя (Maja) |
488 |
44,1 |
13,6 |
12,4 |
554 |
14,5 |
Зелений краб (Carcinus) |
531 |
19,5 |
13,3 |
12,3 |
557 |
16,5 |
Омар (Nephrops) |
541 |
9,3 |
И,9 |
7,8 |
552 |
19,8 |
Міксина (Мухіпе) |
537 |
18,0 |
5,9 |
9,1 |
542 |
6,3 |
У голкошкірих не виявлено істотної регуляції вмісту іонів у рідинах тіла. Кишковопорожнинні регулюють концентрацію тільки де яких іонів, зокрема медуза Aurelia — сульфату. У членистоногих, які здатні швидко рухатися, рівень магнію у крові низький (у краба Pachygrapsus, омара Nephrops), а в тих, які повільно рухаються (наприклад, краб Maja), концентрація магнію висока. Разом з тим у каракатиці Sepia, гребінця Pecten, які рухаються швидко, вміст магнію у рідинах тіла теж високий. Отже, не існує прямої залежності між вмістом магнію в рідинах тіла, який здатний пригнічувати нервово-м’язове передавання сигналів і швидкістю руху морських безхребетних.
Якщо морських безхребетних перенести в розведену морську воду, то в деяких осморегуляторів (морські зірки, устриці тощо) вміст іонів у рідинах тіла стає таким самим, як і у воді. Проте це спостерігається тільки тоді, коли морська вода розведена в певних межах. Так, краб Carcinus не витримує в солонуватій воді, якщо концентрація солей у ній становить менш як 30 % від морської води. Водночас краб Eriocheir витримує значно більші розведення і здатний проникати в прісні водоймища, але для розмноження він повертається в море. Голкошкірі і головоногі молюски (кальмари і восьминоги) у прісній воді існувати не можуть.
Креветки Palaemonetes і Leander у морській воді гіпотонічні — рідини їхнього тіла мають меншу концентрацію солей, ніж довкілля. Щоб це відбувалося, необхідна активна осморегуляція. Така гіпорегуляція в цих креветок здійснюється, напевно, тому що вони раніше мешкали в прісній воді, а в море переселилися згодом. Саме тому вони зберегли концентрацію солей у рідинах тіла нижчою, ніж у морській воді.
Вода деяких солоних водойм містить більше солей, ніж морська. За цих умов особливе значення має гіпорегуляція вмісту солей організмами. Рачок Artemia здатний існувати у воді, яка в 10 разів менш солона, ніж морська, а також у такій, де відбувається кристалізація солі (300 г солі віл води). У концентрованому сольовому розчині цей рачок гіпотонічний завдяки активній осморегуляції.
Личинки деяких комарів, зокрема Aedes campestris, здатні існувати як у прісній, так і солоній воді, оскільки в них теж наявна система активної регуляції солей.
Водні хребетні тварини. Морських хребетних за саморегуляцією можна розподілити на дві групи: 1) ті, у яких концентрація солей у рідинах тіла така, як і в морській воді, або трохи вища (міксини, пластинчатозяброві, целакант Latimeria, крабоїдна жаба та ін.); 2) концентрація солей у рідинах тіла майже втричі нижча, ніж у морській воді (міноги, кісткові риби тощо).
Круглороті мають форму, подібну до вугрів. До них належать міноги й міксини. Міноги здатні існувати в морській і прісній воді, а міксини — тільки в морській. У міксин концентрація солей у рідинах тіла подібна до їхньої кількості в морській воді, а натрію навіть трохи більше. Разом з тим міксини здатні до іонної регуляції. В інших морських хребетних концентрація солей нижча, ніж у довкіллі. Навіть морська мінога Petromyzon marinus належить до анадромних (гр. anos — вгору і dromos — біг) організмів, які піднімаються для нересту з моря в річки.
Майже всі пластинчатозяброві (акули, скати) — морські тварини. Вони підтримують концентрацію солей у рідинах тіла на рівні приблизно втричі нижчому, ніж у морській воді, але при цьому зберігають осмотичну рівновагу. Це досягається шляхом надходження в рідини тіла органічних речовин, головним чином сечовини, унаслідок чого загальна осмотична концентрація крові дорівнює або навіть дещо вища, ніж концентрація морської води.
Сечовина — це кінцевий продукт білкового обміну у ссавців і деяких інших хребетних. У ссавців вона виводиться із сечею, але в акули нирки реабсорбують сечовину, яка залишається в крові.
Крім сечовини, у пластинчатозябрових у кров може потрапляти оксид триметилаланіну (рис. 8.2), який трапляється в багатьох морських тварин.
Пластинчатозяброві, незважаючи на здатність підтримувати ізоосмотичність, здатні регулювати іонний склад рідин тіла. Так, концентрація натрію підтримується приблизно вдвічі нижчою, ніж у морській воді. Надлишок натрію виводиться через нирки й ректальну залозу. Вона відкривається в пряму кишку й виділяє рідину з високою концентрацією, більшою, ніж у води, натрію та хлору.
Деякі пластинчатозяброві здатні заходити в прісну воду, а також постійно жити в ній. У таких тварин у крові низький рівень розчинених речовин, навіть менший, ніж у ссавців.
Морські кісткові риби, як уже зазначалося, здатні підтримувати концентрацію солей утричі нижчою, ніж у морській воді. Ці риби гіпоосмотичні. Для відшкодування осмотичної втрати вони споживають морську воду, шо призводить до всмоктування великої кількості солей. Крім цього, морська вода потрапляє в організм кісткових риб через зябра, а деяка кількість — через поверхню тіла. Виведення надлишків солей відбувається в основному активним шляхом через зябра, оскільки в цих риб сеча зазвичай більш розбавлена, ніж рідини тіла. Однак нирки беруть участь у виділенні іонів магнію й сульфату (Mg2+, SO4-2), які становлять приблизно десяту частку всіх солей морської води. Ці іони не виділяються через зябра.

Рис. 8.2. Структура сечовини й оксиду триметилаланіну
У прісноводних кісткових риб концентрація солей вища, ніж у навколишній воді. Наслідком цього може бути осмотичний приплив води. Надлишок води виводиться у вигляді сечі. Розчинні у воді речовини поглинаються активним способом через зябра і, очевидно, незначна кількість — через поверхню тіла.
Кісткові риби мають обмежену здатність переходити з прісної води в морську й назад. Проте деякі з них, як уже було сказано, мають цю властивість — міноги, вугри, лососі. У них при переході з прісної в морську воду й назад змінюється напрям не тільки осмотичного припливу води, а й активного транспорту солей у зябрах.
Більшість земноводних (амфібій) — напівводяні тварини. Вони відкладають яйця у воду, і їхні личинки живуть у воді та дихають зябрами. Дорослі особини переходять на легеневе дихання. Однак деякі хвостаті амфібії і в дорослому стані продовжують дихати зябрами й залишаються водними тваринами.
Майже всі амфібії — прісноводні тварини. У дорослих особин головним органом осморетуляцїї є шкіра. Коли вони перебувають у воді, відбувається осмотичне поглинання води, надлишки якої виводяться в основному із сечею. Втрати певної кількості солей із водою, яка виводиться з організму, відшкодовуються їх активним транспортом через шкіру.
Одна із жаб, а саме крабоїдна Rana cancerivora, мешкає в морській воді. Для підтримування концентрації солей у рідинах тіла вона використовує той самий спосіб, що і пластинчатозяброві, — у рідини тіла виводиться велика кількість сечовини, необхідна для нормального скорочення м’язів. На відміну від акул, у яких відбувається реабсорбція сечовини в нирках, крабоїдна жаба нагромаджує сечовину внаслідок зменшення об’єму сечі, коли вона перебуває в морській воді. У пуголовок крабоїдної жаби спосіб осморегуляцїї подібний до кісткових риб — виведення надлишків солей відбувається активним шляхом через зябра.
Запліднення яєць і метаморфози крабоїдної жаби відбуваються у воді з низькою концентрацією солей, а саме в прибережних водах, які насичуються прісною річковою й дощовою водою.
Наземні тварини з вологою шкірою. Випаровування води з поверхні тіла зростає з підвищенням температури, у сухому повітрі воно відбувається швидше, ніж у вологому. Дощовий черв’як риє ходи у ґрунті, де повітря насичене вологою і де його тіло стикається із частинками грунту, що вкриті тонкою плівкою води.
Дорослі земноводні, які мають вологу шкіру, мешкають поблизу води й у вологих місцях, де випарювання невелике. Деякі з них (зокрема, жаба Chiroleptes) пристосувалися до життя навіть у пустелях. Вони ховаються в норах, де проводять у сплячці весь період засухи. Під час дощів ці жаби виходять із норок, відновлюють нормальний вміст води в організмі, відкладають яйця в утворені тимчасові водойми. Пуголовки проходять метаморфоз до того, як ці водойми пересохнуть. Коли пустельна жаба знову впадає у сплячку, її сечовий міхур наповнюється розведеною сечею, яка слугує основним запасом води й постійно витрачається під час сплячки.
Пустельні жаби, які проводять період засухи в норах, подібні в цьому аспекті до двоякодихаючих риб, які зариваються в мул.
Деякі жаби не потребують вологого середовища. У шкірі Phyllomedusa містяться особливі залози, які виділяють віскоподібний секрет, що, очевидно, робить шкіру водонепроникною. У Chiro- mantis таких залоз немає, але шкіра цієї жаби теж непроникна для води завдяки особливій будові.
Слимаки також мають вологу шкіру, з якої може випаровуватися вода. Голі слимаки мешкають у вологому середовищі, їхня активність найбільша вночі після дощу. Наземний слимак, який перебуває в неактивному стані, захищається від випаровування води, ховаючись у свою раковину, і закриває отвір висохлою мембраною зі слизу (епіфрагмою). Завдяки цьому деякі слимаки (зокрема, Sphincterochila) здатні мешкати в пустелях. Протягом засушливого сезону слимаки перебувають у стані сплячки. З настанням дощів вони починають вести активний спосіб життя.
Членистоногі. Комахи й павукоподібні (павуки, кліщі, скорпіони тощо) — переважно наземні тварини. Тільки деякі з них вторинно переселилися в прісну воду. Наземними також серед членистоногих є двопарноногі (Diplopoda) та губоногі (Chilopoda). На відміну від членистоногих, більшість ракоподібних — водні тварини, як і меростомові (Xiphosura), які представлені мечохвостом Limulus.
Більшість ракоподібних мешкає у воді, але деякі з них належать до наземних тварин. Такі наземні ракоподібні живуть переважно у вологих місцях і поряд із водою. Водночас деякі мокриці здатні мешкати в пустелях.
Наземні й напівназемні краби здатні до гіпер- і гіпорегуляції водного режиму. Наземні краби Cardisoma і Gecarcinus здатні вбирати воду з піску навіть у разі відсутності її видимої кількості. Деякі з суходольних крабів здатні занурюватись у морську воду (зокрема, Ocypode rotundata). Краби Uca риють глибокі нори на березі біля або вище верхньої межі припливу, але вони часто повертаються у воду.
Рівноногі рачки (мокриці) найчастіше мешкають у вологій місцевості. Інколи вони можуть стикатися із сухим повітрям, наслідком чого є збільшення випаровування води. Для її поповнення вони харчуються, як правило, вологими рослинними залишками. Ці мокриці здатні також пити воду і всмоктувати її через анус. Крім цього, кутикула мокриць, як правило, проникна для води.
У пустелі мешкає мокриця Hemilepistus, яка дихає зябрами. Вона викопує вертикальні нірки завглибшки до ЗО см, у яких проводить спекотні години доби. У цих нірках вологість досягає 95 %. Кутикула Hemilepistus дещо менш проникна для води, ніж в інших мокриць, що зменшує випаровування води через поверхню тіла.
Комахи й павукоподібні нестачу води поповнюють під час пиття, з їжею, у процесі поглинання через поверхню тіла з води або повітря, використовують також воду метаболічних процесів окиснення.
Надходження води до організму тварин відбувається в основному шляхом пиття. Однак у сухих місцях мешкання інколи вільної води не буває. Тоді тварини повинні отримувати воду іншим шляхом. Один з них — це вода в їжі. Комахи, які харчуються певними частинами рослин (листям, плодами та ін.), отримують достатню кількість води з їжею. За надлишку води в їжі вона виводиться з екскрементами.
Комахи, які мешкають у дуже сухому середовищі (наприклад, борошняний черв), вилучають воду з вмісту прямої кишки, поки фекальні кульки не стануть зовсім сухими.
У деяких комах (наприклад, тарганів) екскреторні продукти не виводяться із сечею, а зберігаються в тілі. У цьому разі вода не використовується на їх виділення. Один з таких підходів, це депонування в тілі сечової кислоти, яка малорозчинна у воді. У деяких комах вона відкладається в жировому тілі, в інших — у кутикулі. З кутикули сечова кислота, очевидно, вилучається, а в жировому тілі вона може бути джерелом метаболічного нетрогену.
У водних комах вода потрапляє в організм в основному через поверхню тіла. Наземні комахи (зокрема, пустельний тарган Arenivaga) здатні поглинати воду з атмосферного повітря.
Тварини, які мешкають у безводному середовищі, отримують воду, яка утворюється в організмі під час окиснення органічних спо
лук, —метаболічну воду. Кількість води, яка утворюється в процесах окиснення деяких органічних сполук, наведено в табл. 8.9.
Таблиця 8.9
КІЛЬКІСТЬ ВОДИ, ЯКА УТВОРЮЄТЬСЯ В ОРГАНІЗМІ ПІД ЧАС ОКИСНЕННЯ ДЕЯКИХ ОРГАНІЧНИХ СПОЛУК
Речовина |
Утворення води, гН2О/г речовини |
Жир |
1,07 |
Глюкоза |
0,60 |
Крохмаль |
0,56 |
Білок: |
|
при виділенні сечовини |
0,39 |
при виділенні сечової кислоти |
0,50 |
Плазуни (рептилії). Життя крокодилів назавжди пов’язане з водою, а змій, ящірок і черепах — в основному із суходолом, хоча серед них є водні і напівводні тварини. Усі водні плазуни дихають легенями. Шкіра в плазунів суха, може мати лусочки. У них випаровування води може відбуватися з вологої поверхні дихальних шляхів, а також у більшості цих випадків — через шкіру. Так, наприклад, у вужа Natrix майже 90 % усієї води випаровується з поверхні тіла, а в пустельної ящірки Saumromalus — до 65 %.
Крім втрат води внаслідок випаровування, вона виводиться з організму також із сечею. У більшості плазунів головним кінцевим продуктом білкового обміну є сечова кислота, яка є малорозчинною, тож для її виведення необхідно дуже мало води. Основним джерелом води для наземних плазунів є їжа й метаболічна вода процесів окиснення. Морські плазуни мешкають у збагаченій солями воді, значна частина їхньої їжі містить високі концентрації солі. Морські змії зовсім не зв’язані з суходолом. У морських черепах тільки самки виходять на узбережжя для відкладання яєць. Крокодили, мабуть, не мають дійсно морських представників. Крокодил Crocodylus porosus, який мешкає в солоній воді, харчується переважно рибою й не може довго існувати в справжній морській воді.
Нирки морських плазунів не здатні виводити надлишкову кількість солей. Це робиться спеціальними залозами, що виділяють солі, розміщені в голові. Вони секретують рідину, яка сильно збагачена в основному натрієм і хлором. Ці залози, на відміну від нирок, функціонують не безперервно, а тільки реагують на солоне навантаження.
У морських ящірок залози, що виділяють солі, відкриваються в передню частину носової порожнини і за різкого видиху рідина викидається з ніздрів у вигляді дрібних бризок.
Морські черепахи містять залози, що виділяють солі в орбітах обох очей, а в морських змій їхні протоки відкриваються в ротову порожнину, звідки їхній секрет виводиться назовні.
Наземні плазуни теж містять носові залози, які виділяють секрет із дуже високим вмістом солей. Особливістю залоз, що виділяють солі наземних плазунів, є те, що їхнє виділення залежить від характеру сольового навантаження: якщо є надлишок натрію, то й натрій виділятиметься назовні, а якщо калію, то калій буде в секреті залоз, які виділяють солі.
Птахи. Основними джерелами води для наземних птахів є питна вода, з їжею, а також метаболічна вода процесів окиснення. Виведення надлишку солей відбувається передусім із сечею. Вони мають також солевивідні залози, але невеликі за розміром порівняно з морськими птахами.
Немало птахів живе на морі або над ним. Багато птахів мешкає на березі. У всіх морських птахів добре розвинуті парні солевиділяючі залози, з’єднані з носовою порожниною.
Залози птахів, що виділяють солі, найчастіше розміщені на верхній частині черепа, над орбітою нижнього ока, у неглибокій западині кістки. Ці залози функціонують тільки у відповідь на надлишок солі в організмі, на відміну від безперервної роботи нирок. Вміст солі в секреті залоз, що виділяють солі, морських птахів неоднаковий у різних видів. Так, концентрація натрію в секреті морських птахів коливається в межах 400 — 1100 ммопь/л. Рідина, яка секретується залозами, що виділяють солі морських птахів, містить натрій і мало калію.
Ссавці. У пустелях вільна вода, крім оазисів, з’являється під час дощів. У пустельній місцевості мешкають дрібні ссавці, а саме гризуни. Основний спосіб надходження води в їхній організм —- з їжею. Роса для пустельних тварин як джерело води відіграє мінімальну роль. Водночас деякі з гризунів пустелі харчуються сухим насінням та іншим сухим рослинним матеріалом і отримують вільну воду в мінімальній кількості. Основне джерело води для них — метаболічна вода, яка утворюється в окисних процесах. Крім цього такі тварини (типовим представником є кенгуровий щур) здатні зводити до мінімуму втрати води із сечею й калом. Повітря, яке видихається під час проходження через ніс, у них охолоджується, що знижує випаровування води через органи дихання.
Представники рядів ластоногі, кити й сирени (ламантин і дюгонь) є морськими тваринами в тому розумінні, що майже все своє життя проводять у морі. Ластоногі ненадовго повертаються на суходіл для народження й вигодовування малят, а кити та сирени навіть народжують нащадків у воді.
їжа морських ссавців дуже різноманітна за вмістом солей. Так, тюлені, які харчуються переважно рибою, споживають мало солі, але багато білка. Разом з тим антарктичний крабоїдний тюлень і морж (харчуються в основному двостулковими молюсками) з їжею вживають велику кількість солі. Беззубі кити споживають солевмісних планктонних безхребетних. Якщо з їжею заковтується морська вода, то це ще збільшує сольове навантаження. Ламантини й дюгони харчуються морськими рослинами, які містять велику кількість солей.
Основний шлях виведення надлишку солей у морських ссавців — це виділення сечі, яка концентрованіша, ніж морська вода.
Самки ссавців вигодовують малят молоком, для якого необхідна велика кількість води. Зменшити таку втрату води можна концентруванням молока. У молоці тюленів і китів дуже високий вміст жиру й концентрація білків вища, ніж у молоці корови. Дитинчати цих тварин повинні швидко розвиватися й отримувати велику кількість жиру для відкладання його під шкірою, де він слугує теплоізоляцією. Разом з тим високий вміст жиру в молоці тюленів можна розглядати у зв’язку з обмеженням водних ресурсів матері. Тюлені вигодовують своїх малят за мінімальних затрат води.
Відомо, що для людини морська вода токсична і прискорює зневоднення організму при її споживанні. Нирки людини не здатні вилучати кількості солей, які містяться в морській воді: максимальна концентрація солей у сечі людини нижча, ніж у морській воді. Крім цього, зневодненню організму сприяє велика кількість магнію і сульфату в морській воді, які викликають пронос і, отже, збільшують втрату води.