ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ 8. СТІЙКІСТЬ ТА АДАПТАЦІЯ РОСЛИН І ТВАРИН
8.3. Стійкість та адаптація тварин до умов існування
8.3.6. Біолюмінесценція
Здатність до свічення значно поширена серед бактерій, грибів, водоростей і тварин. Переважна більшість тварин, які світяться, належить до морських глибоководних, прибережних і планктонових організмів. Проте світяться також жуки-світляки, слимаки, деякі личинки двокрилих тощо.
Люмінесценція (лат. lumen — свічення і суфікс escent — слабка дія) у тварин є результатом хемілюмінесценції, тобто зумовлена хімічними реакціями, у процесі яких окиснення субстрату приводить до свічення. За внутрішньоклітинної люмінесценції такі реакції відбуваються в розміщених у цитозолі гранулах клітин тканин, які люмінесціюють. Позаклітинна люмінесценція відбувається у світлогенеруючому органі, який становить собою одноклітинні або багатоклітинні залози, що виділяють секрет назовні. Інколи в цих залозах існують два типи клітин, які містять окремо субстрат (люциферин) і фермент (люциферазу), що беруть участь у процесі генерації світла.
Люмінесценція організмів має певне значення для їхнього виживання і з’явилась у процесі еволюції, оскільки для цього розвинулися спеціальні клітини та органи. Крім того, це явище може бути частиною загальної здатності організмів відповідати на подразнення.
Явище люмінесценції виконує, перш за все, захисну функцію — слугує для попередження, відлякування та відвернення хижаків. Воно допомагає тварині сховатись у хмарці, що світиться, як .це має місце в глибоководної морської креветки Acanihophyra й кальмара Heteroteuthis. Можливо також, що стимуляція свічення однієї особини слугує попередженням про небезпеку іншим особинам і дозволяє їм уникати небезпеки.
Ще однією з функцій люмінесценції тварин є сигнальна для зустрічі самців і самок у шлюбний період. Типовим прикладом є приваблення самців самками жуків-світляків. У багатьох їх видів самки, іноді навіть безкрилі, сидять у траві й подають світлові сигнали самцям, які літають навколо них. Самка відповідає на сигнали свічення самця тільки свого виду цілком певним чином — спалахами певної частоти, кольору, інтенсивності, тривалістю та їх кількістю.
Явище люмінесценції тварин зумовлює агресивну мімікрію, коли самки одних видів приваблюють самців інших видів, яких поїдають. Таке спостерігається, наприклад, у самок світляків Photuris, які полюють на інших видів самців цієї родини.
У найпростіших люмінесцентні гранули розсіяні по всій цитоплазмі, а у вищих тварин фотогенеруючі клітини або органи розміщені в певному місці. У головоногих молюсків, ракоподібних, риб і комах клітини, які світяться, становлять тільки частину органа, який містить також шари клітин (певні «світофільтри»), які поглинають та відбивають світло і заломлюють світло тільця, нервові закінчення.
Іноді свічення організмів зумовлене не власною люмінесценцією, а свіченням наявних в організмі та на його поверхні бактерій, які люмінесціюють.
Поширеність біолюмінесценції серед тварин. Серед найпростіших існує багато морських видів, які люмінесціюють, зокрема радіоляції та динофлагеляти. В останніх відносно добре вивчені Gonyaulax та Noctiluca. Гранули, які люмінесціюють, локалізовані в мікросомах, але найбільше їх на периферії клітин. Вони світяться у відповідь на механічне або електричне подразнення.
У Gonyaulax, Peridinium і Pyrocystis люмінесценція виникає як у відповідь на збудження, так і спонтанно. Залежно від часу доби інтенсивність їхньої люмінесценції може змінюватися.
Серед кишковопорожнинних до люмінесценції здатні багато видів поліпів, медуз, сифанофорів, актиній. Це явище в них виникає у відповідь на подразнення. Першою відповідає свіченням ділянка,
яка була безпосередньо механічно подразнена, а потім від неї поширюється хвиля свічення. У деяких кишковопорожнинних свічення може мати позаклітинну природу.
Люмінесціюють, очевидно, усі реброплави (тип Ctenopohora) у відповідь на подразнення. Світло інактивує це свічення.
Серед немертин здатні до люмінесценції тільки Emplectonema randai. Фотогенеруючі клітини розміщені по всьому тілу. Місцеве тактильне подразнення Викликає локальну відповідь. До люмінесценції серед кільчастих червів здатні морські поліхети та Наземні олігохети.
Деякі дощові черви, наприклад Eisenia submontana, при подразненні викидають через ротовий чи анальний отвір, через дорсальні пОрй слиз, який світиться. У поліхети Chaetopterus за подразнення починає світитися більша частина її поверхні. Фотогенеруючі клітини розміщені в гіподермі разом зі слизовими клітинами, які разом виділяються назовні. Свічення в кільчастих червів регулюється нервовою системою. Подразнення частини їхнього тіла викликає хвилю світла, яка проходить вгору і вниз від точки збудження.
Серед членистоногих до люмінесценції здатні в основному ракоподібні та комахи, а також деякі багатоніжки й павукоподібні. Саме вивчення свічення рачків роду Cypridina і жуків-світляків дало основні відомості про хімічні процеси біолюмінесценції. У Cypridina світлогенеруючий орган становить залозу, розміщену поблизу ротового отвору. Деякі з секреторних клітин містять субстрат, а інші — фермент.
В евфаузидій і креветок фотогенеруючі органи розміщені по всій поверхні тіла й регулюються нервовою системою. Глибоководна креветка Acanthophyra purpurea, крім типових фотогенеруючих органів біля ротового отвору, містить також залози, з яких викидається речовина, що світиться. Це дає можливість креветці, яка оточує себе хмаркою, що світиться, ховатися від ворогів.
Серед комах світяться представники лише кількох рядів. Здатними до люмінесценції є кілька видів багатохвосток Collembola, личинки грибного комара Ceratoplanus, личинки й дорослі особини грибного комара Bolitophila. Свічення в них генерується в мальпігієвих канальцях.
Найбільш вивченими комахами, які здатні до люмінесценції, є Coleoptera, зокрема жуки-світляки Lampyridae і лускунчики Elateridae. У них фотогенеруючий орган розміщений у задній ділянці черевця. Він складається із шару фотогенеруючих клітин і дорзального, або внутрішнього, шару клітин, які відбивають світло. Свічення комах регулюється нервовою системою.
У південноамериканського лускунчика Pyrophorus пара органів, які люмінесціюють, зеленуватого кольору розміщені на передній частині грудини, а оранжево-жовтий орган — на черевній поверхні
першого черевного сегмента. Голова південноафриканського «залізничного черва» Phrixothrix світиться червоним світлом, а по всьому тілу сегментарно розміщені плями зеленого кольору, які люмінесціюють. Свічення в цієї комахи відбувається у відповідь на подразнення. Водночас північноамериканський рід комах Phengodes світиться безперервно, і в них немає органа свічення на голові.
Деякі види багатоніжок виділяють слиз, який світиться подібно до такого в кільчастих червів. Описано свічення також морського павука
Серед молюсків найбільшу здатність до люмінесценції мають головоногі. У багатьох із них, наприклад у кальмара Loligo, орган свічення розташований назовні й містить бактерії, які світяться. Сам же кальмар Loligo не містить органів, які люмінесціюють. Разом з тим у більшості молюсків наявні органи, здатні до люмінесценції. Так, у кальмара Heteroteuthis непарний орган, здатний до люмінесценції, відкривається в порожнину мантії, куди через сифон під час подразнення викидається хмарка, що світиться. У кальмара Watseniaфотогенеруючі органи розташовані по всьому тілу, зокрема по три з них містяться на кожній кінцівці.
У голкошкірих органи, які люмінесціюють, містять одноклітинні фотогенеруючі органи, розсіяні по всьому тілу. Люмінесценція в них виникає у відповідь на будь-яке подразнення.
До люмінесценції здатні також нижчі хордові та риби. У покривників (наприклад, Pyrosoma) під час подразнення хвиля світла поширюється по всій колонії. Деякі види Balanoglossus секретують слиз, який світиться.
Багато видів, здатних до люмінесценції, виявлено серед акулових і кісткових риб, особливо глибоководних. Добре розвинуті фотогенеруючі органи голови притаманні Astronesthes та Stomias.
У деяких риб (Photoblepharon, Anomalops, Physiculus, Leiognathus, Monocentrus та ін.) свічення генерується бактеріями, які містяться в особливих органах і виділяються назовні під час подразнення.
Хімічні механізми люмінесценції тварин. Біолюмінесценція, як уже зазначалося, є хемілюмінесценцією. З’ясовано, що це явище виникає в процесі реакції окиснення субстрату, який називають люциферином (лат. lucis — світло і ferre — носити), молекулярним оксигеном за участю ферменту люциферази. Цей процес можна представити рівнянням (рис. 8.7).
Різні організми містять специфічні субстрати й ферменти, але механізм виникнення в них явища люмінесценції схожий. Тому терміни «люциферин» і «люцифераза» використовують для позначення субстрату й ферменту в хімічних реакціях, які супроводжуються люмінесценцією.
![]()
Рис. 8.7. Рівняння реакції субстрату й ферменту, яке супроводжується люмінесценцією:
S — субстрат (люциферин); Е — фермент (люцифераза); E - S* — фермент-субстратний комплекс в електроннозбудженому стані; hv — квант світла (h — стала Планка, v—частота)
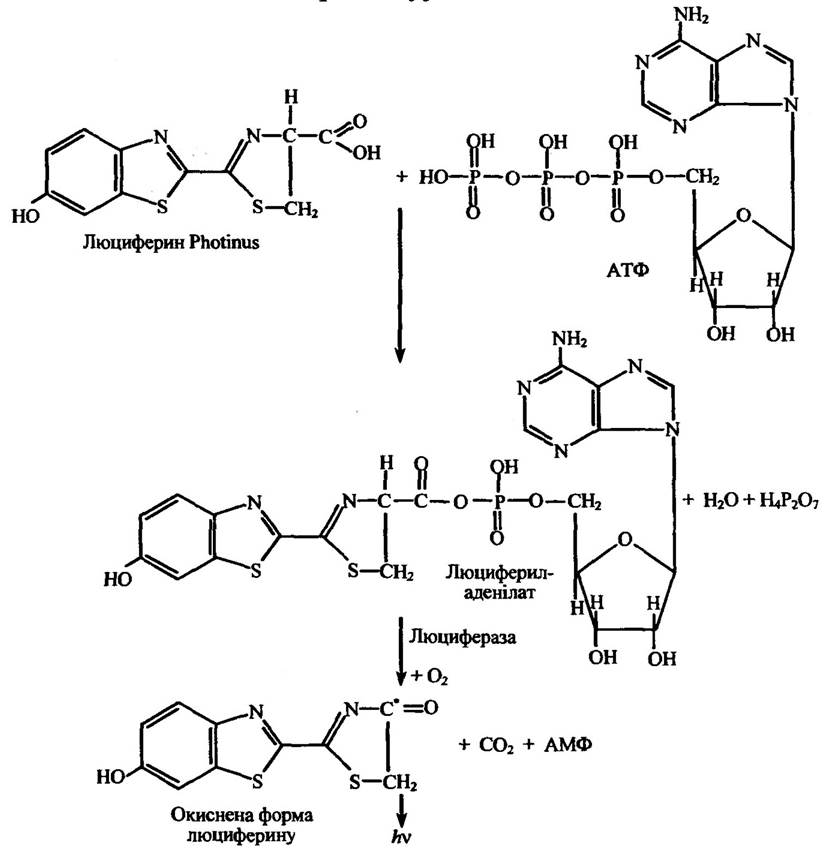
Рис. 8.8. Реакція взаємодії люциферину жука-світляка (Photinus) з АТФ у присутності люциферази,
яка супроводжується люмінесценцією
Примітка. Позначка (*) означає, що даний атом перебуває в електронно збудженому стані.
Розглянемо перебіг реакцій люциферину й люциферази в жука-світляка Photinus, а також рачка Cypridina.
У жука-світляка реакція, яка супроводжується люмінесценцією, відбувається внутрішньоклітинно. Його люциферин активується в процесі реакції з АТФ з утворенням люцифериладенілату в електронно збудженому стані, повернення якого в основний стан супроводжується свіченням у червоній області спектра (X = 560 нм). Схематично це ілюструє рис. 8.8.
Різниця в спектрах люмінесценції різних видів жуків-світляків зумовлена в основному відмінностями структури й конформації люциферази.
Реакцію взаємодії люциферину й люциферази рачка Cypridina, яка спряжена зі свіченням, наведено на рис. 8.9.
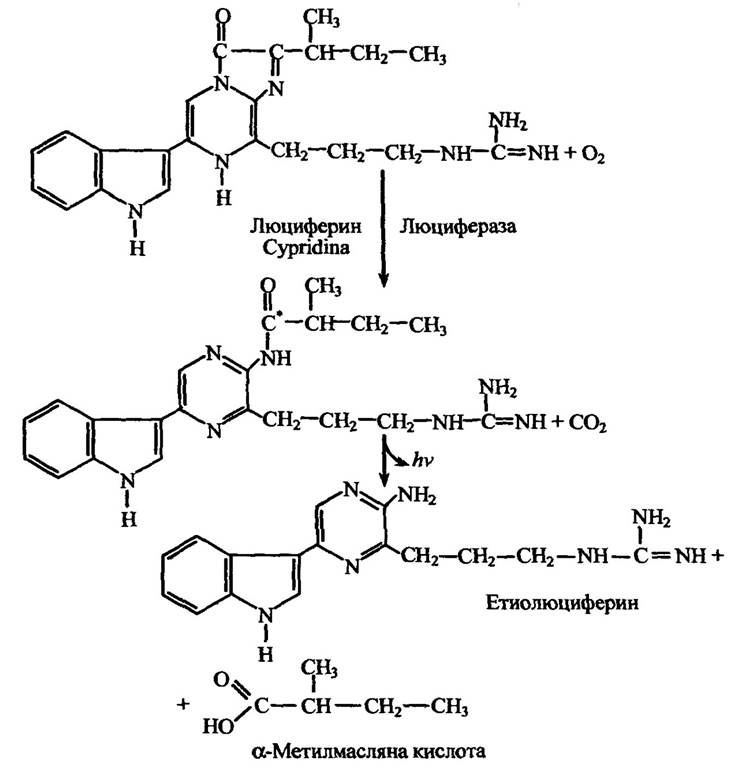
Рис. 8.9. Реакція взаємодії люциферину Cypridhia у присутності люциферази, яка супроводжується люмінесценцією
Примітка. Позначка (*) означає, що даний атом перебуває в електронно збудженому стані.
Докладно вивчено біохімічні процеси, покладені в основу люмінесценції в деяких видів кишковопорожнинних. Так, наприклад, у коралів Renilla цей процес розпадається на дві стадії (рис. 8.10).
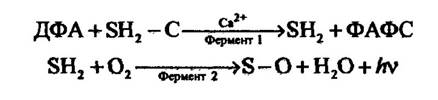
Рис. 8.10. Рівняння реакцій субстратів і ферментів у Renilla, які супроводжуються люмінесценцією:
ДФА — 3', 5-дифосфоаденозин; SH2 - С — люциферин; SH2 — люциферин з вилученою групою; ФАФС — фосфоаденозил-фосфорилсульфат
На першій стадії активація люциферину відбувається при вилученні однієї з груп, яка ідентифікована як сульфат. Ця реакція стимулюється 3'5-дифосфоаденозином. Наслідком її є утворення активованого люциферину й фосфоаденозилфосфорилсульфату. Активований люциферин за структурою подібний до люциферину рачка Cypridina. За його окиснення, механізм якого нагадує окиснення люциферину Cypridina, відбувається явище люмінесценції.
У медуз Aeguorea і Halistaurea, а також деяких інших видів медуз виявлено білок (фотобілок), у якого при взаємодії з іонами кальцію за наявності оксигену виникає явище люмінесценції. Цей білок, мабуть, є проміжним продуктом ланцюга реакцій, які подібні до тих, що описані для корала Renilla. На останній стадії перетворення фотобілок взаємодіє з оксигеном за наявності іонів кальцію.
У деяких кишковопорожнинних при люмінесценції відбувається перенесення енергії: спочатку в реакції утворюється електроннозбуджений продукт, який потім передає енергію іншій речовині, за повернення якої в основний стан після збудження виникає явище люмінесценції (рис. 8.11).
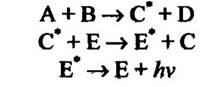
Рис. 8.11. Рівняння реакцій люмінесценції з перенесенням енергії в кишковопорожнинних
Примітка. Позначка (*) означає, що даний атом перебуває в електронно збудженому стані
Система люмінесценції кільчастого черв’яка Chaetopterus містить фотобілок, два кофактори (один з яких подібний до нуклеопротеїну, а інший — ліпіду), пероксиди (Н2О2 або органічні пероксиди), оксиген і ферум.
У риб, які здатні до власної люмінесценції, а не за рахунок симбіотичних бактерій (зокрема, Apogon і Parapriacanthus), утворення люциферину, можливо, викликане тим, що ці риби харчуються рачками Cypridina. Разом з тим люцифераза цих риб не подібна до лю циферази Cypridina.
У бактерій спостерігається внутрішньоклітинне безперервне свічення, в основу якого покладено, наприклад, окиснення відновленого рибофлавін-5'-фосфату (флавінмоноклеотиду) (рис. 8.12).
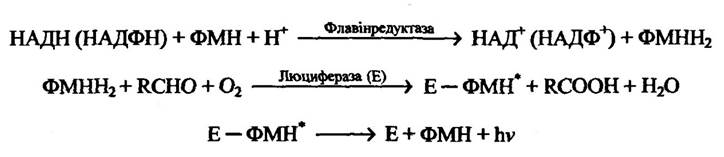
Рис. 8.12. Реакція окиснення рибофлавін-5'-фосфату, що супроводжується біолюмінісценцією бактерій
Примітка. Позначка (*) означає, що даний атом перебуває в електронно збудженому стані
Люмінесценція грибів охоплює принаймні дві стадії: 1) відновлення дегідро- або оксилюциферину за участі НАДН (НАДФН) з ферментом; 2) окиснення відновленого продукту оксигеном у присутності ферменту люциферази з випромінюванням світла.
Отже, за всіх відмінностей у способах перебігу хімічних реакцій, які супроводжуються люмінесценцією, загальним є окиснення люциферину за участю люциферази з утворенням електронно збудженого стану продукту, за повернення якого в основний стан випромінюється світло.