ЕКОЛОГІЧНА БІОХІМІЯ - Навчальний посібник - В. М. Ісаєнко 2005
Розділ І. ВІРУСНІ ІНФЕКЦІЇ ТА РЕАКЦІЯ НА НИХ ОРГАНІЗМІВ (загальні уявлення)
1.3. Віруси рослин
Віруси рослин, або більш уживаний термін інфікують як нижчі, так і вищі рослини. Зауважимо, що саме відкриття вірусів Д. І. Івановським (1892 р.) відбулося під час дослідження вірусу тютюнової мозаїки (ВТМ).
Найкраще досліджені віруси вищих рослин, зокрема покритонасінних, особливо сільськогосподарських культур. На відміну від насіннєвих рослин (голонасінних і, особливо, покритонасінних), віруси спорових рослин (мохоподібних, риніоподібних, псилотоподібних, плавуноподібних і хвощеподібних) вивчено значно менше. Проте ці рослини теж інфікуються фітовірусами. Водночас останнім часом розпочалося дослідження фітовірусів і нижчих рослин, що зумовлено визначенням їхньої ролі в стабільності біорізноманіття. Так, наприклад, у водоростей у більшості випадків виявлено відносно великі поліедричні віруси з чіткими обрисами діаметром 150—250 нм. їх знайдено, зокрема, у клітинах зелених водоростей Oedogonium spp., Uronema gigas тощо, бурої водорості Chorda tomentosa. В ядрах зеленої водорості Coleochaete scutata можуть бути наявні скупчення ізометричного вірусу діаметром 40 нм (ДНК-вмісний вірус). Аналогічні віруси знайдено в клітинах червоної водорості Sirodotia tenuissima. Вірус зеленої водорості Chara corallina містить одноланцюгову РНК і за формою нагадує ВТМ, але більший — його довжина становить 530 нм. Безумовно, перелік досліджених вірусів нижчих рослин значно більший.
Поширення вірусів по рослині. В експериментальних дослідженнях з вищими рослинами показано, що віруси, уведені в рослини шляхом інокуляції, відносно повільно розповсюджуються по непровідній тканині від первинно інфікованих клітин до сусідніх. Так, наприклад, швидкість поширення ВТМ становить приблизно 1 мм за добу. Мабуть, вірус, який потрапив у клітину, спочатку в ній розмножується, а потім проникає в сусідні клітини міжклітинними канальцями або плазмодесмами. Перед тим, як вірус починає рухатися з первинно інфікованої клітини в сусідню, минає декілька годин. З однієї клітини в іншу можуть мігрувати як інтактні віріони, так і вірусна нуклеїнова кислота.
Вірус, який потрапляє в провідну тканину листка із сусідніх паренхімних клітин або безпосередньо вводиться переносниками (зокрема, комахами), рухається спочатку по прожилках, потім по черешку листка і, нарешті, потрапляє в стебло. Узагалі вірус може поширюватися по всій рослині, причому швидкість цього процесу залежить від властивостей як вірусу, так і рослини. Більшість вірусів рослин належать до «необмежених» вірусів, здатних проникати в усі тканини інфікованої рослини. Перенесення вірусів на значні відстані відбувається, головним чином, по флоемі, хоча вони здатні мігрувати й по ксилемі. Перенесення вірусів насінням спостерігається відносно рідко, а пилком — ще рідше. Особливості процесу морфогенезу квітки, мабуть, такі, що вони певною мірою перешкоджають проникненню вірусів у гамети.
Симптоми зараження можуть бути різними й залежать від рослини, тривалості інфекції, штаму вірусу та зовнішніх умов. Наприклад, симптоми захворювання найчіткіше виражені в рослин, вирощених за яскравого освітлення та помірної температури. Вони можуть бути не так помітні за слабкого освітлення та високої температури. Строк від моменту надходження вірусу в клітини рослин до появи симптомів також залежить від штаму вірусу, рослини й умов навколишнього середовища. Для трав’янистих рослин інкубаційний період зазвичай становить кілька днів або тижнів, а для дерев’янистих—він молю бути більше року.
Порушення клітинного метаболізму вірусами. Ступінь ураження вірусною інфекцією рослин визначається реакцією клітин на зараження. Найбільш вираженим її проявом є некротичні ураження. У цьому разі клітини гинуть так швидко, що можуть не встигнути передати вірус сусіднім клітинам. Однак до найпоширеніших проявів реакції рослин на зараження вірусом належать майже безсимптомні інфекції, які виявляються за дуже слабкими симптомами або завдяки тому, що вірус викликає захворювання інших рослин. При цьому інфіковані клітини зберігають здатність до поділу. Одна із реакцій клітин рослин на інфекцію—інтенсивний їх поділ і навіть пухлинна трансформація.
Більша частина порушення обмінних процесів у клітинах рослин за інфекції виникає, мабуть, як наслідок опосередкованих впливів вірусів на метаболічні процеси.
Викликані вірусами зміни метаболізму подібні до тих, які спостерігаються під час старіння рослин. Так, наприклад, при зараженні тютюну вірусом огіркової мозаїки підвищується вміст спирторозчинного нітрогену (амінного, амідного та амонійного), а також фосфору РНК та ДНЮ, тимчасом як концентрація інших органічних сполук фосфору знижується. Інфікування листків тютюну ВТМ мало впливає на вміст загального нітрогену й лише незначно підвищує вміст білкового нітрогену, хоча вірус може містити до 75 % білка листків, тобто синтез вірусних білків відбувається за рахунок нормальних білків листків.
Відомо, що ряд вірусів впливає на функціональну активність хлоропластів і цей вплив дуже специфічний. Наприклад, у період максимальної реплікації вірусу жовтої мозаїки турнепсу в листках спостерігаються тимчасові зміни, які полягають у використанні продуктів фотосинтезу на синтез не цукрів, а амінокислот, а також інших органічних кислот, і в збільшенні активності фосфоенолпіруватрибоксилази й аспартатамінотрансферази. Інші зміни в хлоропластах, вочевидь, менш специфічні і, звичайно, спостерігаються після періоду реплікації вірусу. Вони приводять до зменшення інтенсивності фотосинтетичної фіксації карбону, кількості та активності ферментів (зокрема, рибулозодифосфаткарбоксилази), кількості хлорофілу і вмісту хлоропластних рибосом. Концентрація цитоплазматичних рибосом, навпаки, може не змінюватися.
Листки рослин, уражені вірусами, які викликають пожовтіння, наприклад вірусами жовтяниці цукрового буряку, містять надлишок глюкози, фруктози й сахарози. Первинна причина такого нагромадження цукрів, як вважається, полягає не в блокуванні їх переміщення, а в тому, що в інфікованих рослинах зростає «опір» на шляху переміщення цукрів у черешках і для такої, як раніше, інтенсивності переміщення цукрів з листків у корені необхідний більший градієнт їх концентрацій.
Інший поширений вплив вірусної інфекції проявляється в зниженні як інтенсивності утворення крохмалю в клітинах листків удень, так і швидкості його відпливу з листків уночі.
У разі інфекції, як правило, збільшується інтенсивність процесів дихання рослин. Сильніша стимуляція дихання найчастіше спричиняється тими вірусами або штамами вірусів, у разі зараження якими симптоми хвороби виражені найяскравіше, і ніяких змін дихання не відбувається, якщо симптоми виражені слабко. Посилення дихання в листках Nicotiana, на яких під впливом ВТМ з’являються місцеві некрози, пов’язане головним чином з роз’єднанням дихання та окисного фосфорилювання. Інфікування деякими іншими вірусами може призводити до активації ферментів пентозофосфатного шляху.
За інфікування верхніх молодих здорових листків рослини зараженими нижніми листками спостерігається стійкість до інфекції, що корелює з активацією пероксидази у верхніх листках. Вважають, що активація пероксидази в тканинах призводить до швидшого нагромадження хінонів, які зумовлюють відмирання клітин у спалахах інфекції, і це вповільнює поширення вірусу.
Віруси, що спричиняють утворення виростів (enations) або інші порушення нормального росту, впливають на вміст ростових речовин у рослинах.
Перелік наведених змін метаболізму клітин рослин у разі інфекції далеко не вичерпує всіх відомих. Поширеними проявами інфекції є також збільшення розміру клітин, мацерація тканин, порушення водного режиму внаслідок пошкодження коренів або посиленої транспірації тощо.
Переносники вірусів рослин. Серед багатьох способів поширення вірусів рослин особлива увага належить їх переносникам. Вони захоплюють віруси, зберігають їх у життєздатному стані та вводять у рослину. Наслідком цих процесів є інфікування рослин. Якщо один із цих процесів неможливий, то вірус поширюватися не буде.
Переносниками вірусів рослин є членистоногі, нематоди та гриби (рис. 1.1).
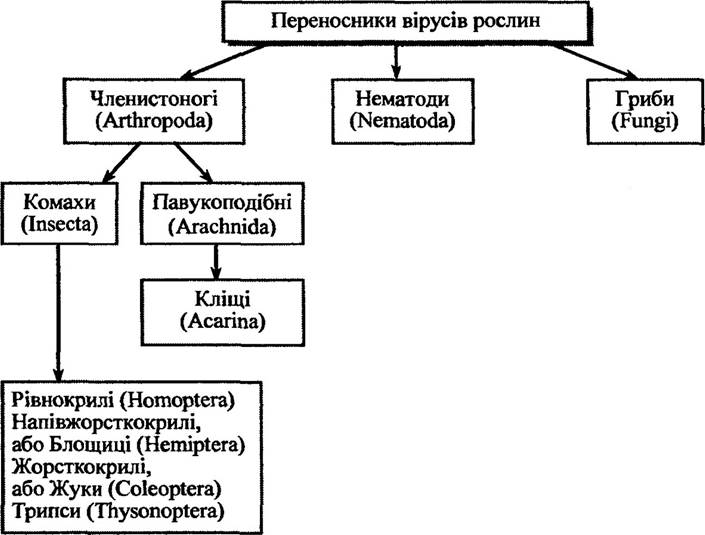
Рис. 1.1. Переносники вірусів рослин (за Рижковою та ін« 2002)
Розглянемо дещо докладніше участь в інфікуванні деяких конкретних переносників вірусів рослин.
Комахи. Рівнокрилих комах налічується близько 25 000 видів. Переносниками вірусів рослин є представники підрядів цикадових (Cicadinea), білокрилок (Aleyrodinea), кокцидій (Coccinea), попелиць (Aphidinea).
Самки цикадових відкладають яйця переважно в стебла. Німфи більшості видів мігрують у грунт, де живляться на підземних частинах рослин. Імаго живляться на надземних частинах рослин і часто перелітають з однієї на іншу, що й зумовлює здатність до передавання фітовірусів.
Найчастіше цикадові переносять віруси рослин родин Caulimoviridae (рід Rice tungo lacilliform-like viruses), Rhabdoviridae (роди Cytorhabdovirus, Nucleoihabdovirus) Sequiviridae(рід Wairavirus), Geminiviridae (роди Mastrevirus, Curtovirus).
Німфи та імаго білокрилок живляться на нижній поверхні листків. Вони переносять віруси рослин з родини Geminiviridae, Closteroviridae, деяких родів Potyviridae (Ipomovirus) та ін. Один з вірусів рослин, які переносяться білокрилками, — гемінні віруси, що викликають в основному пожовтіння, кучерявість, карликовість. Типовими представниками є вірус кучерявості листків томата (Tomato leaf curl virus), вірус золотистої мозаїки томатів (Tomato golden mosaic virus), жовтої мозаїки картоплі (Potato yellow mosaic virus) тощо.
Клостеровіруси, які переносяться білокрилками, викликають передусім пожовтіння між жилками та почервоніння листкових пластинок. Представниками таких вірусів є вірус інфекційного хлорозу латуку (Lettuce infectionus yellow virus). Він вражає латук, цукровий буряк, дині. Ще один представник — вірус інфекційного хлорозу томатів (Tomatoinfectious chlorosis virus).
Виявлено ще багато інших вірусів рослин, які переносяться білокрилками, зокрема крапчастості батату, жовтої кучерявості листків кабачків.
Кокциди (родини Coccidae, Pseudococcidae) переносять в основному віруси представників родів Vitivirus, Badnavirus, Closterovirus.
До найпоширеніших переносників вірусів рослин належать попелиці. У деяких видів частина особин постійно розмножується на первинній рослині-господарі, а в інших — мігрує на вторинну. Такі попелиці, які оселилися на молодих дерев’янистих рослинах, можуть не мігрувати весь сезон, а на старих деревах — переміщуватися на трав’янисті рослини. У деяких видів попелиць міграція на інші рослини є необхідною для завершення життєвого циклу з другого або третього поколінь. Типові представники вірусів рослин, які передаються попелицями, — вірус некротичного пожовтіння салату-латуку (родина вірусів Rhabdoviridae), який розмножується в комасі, вірус жовтої карликовості ячменю (родина Luteoviridae), який не розмножується в переноснику. До вірусів, які передаються попелицями, належить ще ціла низка, зокрема вірус картоплі (вірус скручування листків, Y-вірус, А-вірус, М-вірус, S-вірус).
До комах — переносників вірусів рослин відносять жуків трьох родин — листоїдів (Chrysomelidae), довгоносиків (Curculionidae), сонечок (Coccinellidae). Віруси, що їх переносять жуки, належать до родів Carmovirus (слабкої мозаїки квасолі — Beam mild mosaic vims; зморшкуватості турнепсу — Turnip crinckle vims та ін.), Comovims (мозаїки коров’ячого гороху — Cow pea mosaic vims; мозаїки кабачка — Squash mosaic vims тощо), Bromovirus (крапчастості бобів — Broad bean motile vims та ін.), Sobemovims (південної мозаїки квасолі — Southern bean mosaic vims; жовтої крапчастості рису — Rice yellow motile vims тощо), Tymovims (жовтої мозаїки турнепсу — Turnip yellow mosaic vims; латентний індійської картоплі — Andean potato latent vims та ін.), Machloomovirus (хлоротичної крапчастості кукурудзи — Maize chlorotic motile vims тощо).
Трипси живуть на злакових і осокових, на листках та стеблах чагарників і дерев, на листках і пагонах трав’янистих рослин, під корою засохлих дерев. Значна кількість їх видів живе в суцвіттях, живиться соками зав’язі, нектаром, пилком. Вони переносять віруси родів Tospovims (Bunyaviridae) і Machlomovirus (Tombusviridae). Типові представники вірусів рослин, які передаються трипсами, — хлоротичної плямистості томата (Tomato chlorotic spot vims), некрозу верхівки арахісу (Groundnut but necrosis vims), некрозу стебла хризантеми (Chrysantemum stem but necrosis vims), посрібненої крапчастості дині (Watermelon silver motile vims) та ін.
Кліщі. Серед переносників вірусів рослин істотне значення мають кліщі, які належать до класу павукоподібні, тип членистоногі (рис. 1.1). Це в основному кліщі з родин плескатотілих (Tenuipalpidae), багатощетинкових чотириногих (Phytoptidae) та галових чотириногих (Eriophyidae) ряду Acariformes. Найбільш визначеними в цьому плані є еріофідні кліщі, представником яких є Aceria tulipae.
Кліщі передають різноманітні віруси рослин, зокрема родів Pymovirus (мозаїки житняку — Agropyron mosaic vims; мозаїки ячменю — Hordeum mosaic vims та ін.), Thitimovims(смугастої мозаїки пшениці — Wheat streak mosaic vims тощо), Vepovims (вірус реверсії чорної смородини — Blackcurant reversion assotiate vims), Allexivims (вірус часнику — Carlic virus C; латентний вірус цибулі — Onion mite-borne latent vims та ін.), родин Rhabdoviridae (кільцевої
плямистості кави — Coffee ringspot virus та ін.) та Bromoviridae (кільцевої некротичної плямистості сливи — Prunus necrotic ringspot virus тощо). Показано можливість передавання кліщами вірусів родів Tobamovirus і Potexvirus.
Нематоди. Типовим ектопаразитом є нематоди, які вражають кінчики коренів рослин або клітини, які містяться поблизу. Вони спричиняють зупинку росту рослин або утворення галів незалежно від вірусної інфекції. Водночас нематоди здатні передавати віруси рослин. До таких належать нематоди родин Longidoridae (роди Xiphinema і Longidorus) та Trichodoridae (Trichodorus і Paratrichodorus).
Віруси, які передаються нематодами, є РНК-вмісними і належать до родів Nepovirus (наприклад, кільчастої плямистості тютюну — Tobacco ringspot virus; коротковузля винограду — Grapevine fan leaf virus; чорної кільчастої плямистості томата — Tomato black ring virus; скручування листків вишні — Cherry leafroll virus та ін.) та Tobravirus (погрімковості тютюну — Tobacco rattle virus; раннього побуріння гороху — Pea early-browning virus тощо).
Нематоди родів Xiphinema і Longidorus — відносно великі, їхня довжина S—12 мм. Водночас довжина представників родини Trichodoridae — 0,6—1,2 мм.
Як дорослі особини нематод, так і личинки різних стадій розвитку здатні переносити віруси. Віруси можуть зберігатися в нематоді- переноснику відносно тривалий час — від кількох тижнів до місяців. Дорослі нематоди роду Longidorus, на відміну від Xiphinema, для збереження інфекційної здатності протягом тривалого часу повинні мати постійний доступ до вірусовмісної їжі. Отримання вірусної інфекції нематодами відбувається в інтервалі часу від 15 хв до 1 год після живлення. Нематоди Trichodorus живляться, як правило, на епідермальних клітинах, переходячи через кілька хвилин від одні» до іншої. Тим часом представники роду Xiphinema живляться в одній точці протягом години й більше. Термін, необхідний для зараження рослини вірусом нематодами, коливається від 1 год до кількох діб.
Гриби. Гриби — переносники вірусів рослин є одноклітинними організмами. Вони внутрішньоклітинні паразити, проходять дві стадії — рухливої зооспори та товстостінної спори спокою. Відомо близько 30 вірусів рослин, що передаються грибами класу Chytridiomycetes роду Olpidium та класу Plasmodiophoromycetes родів Polymyxa і Spongospora. Так, наприклад, Olpidium bomovanus передає вірус некрозу огірка (Cucumber necrosis virus), Olpidium brassicae — некрозу тютюну (Tobacco necrosis virus), Polymyxa betae — некротичного пожовтіння жилок буряку (Beet necrotic yellow vein virus), Polymyxa graminis — штрихуватих некрозів рису (Rice stripe necrosis virus), Spongospora subterranea — жовтої плямистості крес-салату (Watercress yellow spot virus) тощо.
Життєвий цикл грибів роду Olpidium відносно простий. Нестатеве розмноження відбувається зооспорами, що мають джгутик. Зооспора з грунтовою водою пересувається до кореня рослини, прикріплюється до нього, втягує джгутик, утворює оболонку цисти та переливає свій вміст у клітину епідермісу рослини. Наслідком цього є утворення талому, який у процесі розвитку стає багатоядерним і перетворюється в зооспорангій. Зооспори виходять через проколену клітинну стінку. Увесь цикл займає 5—10 днів.
За статевого розмноження грибів роду Olpidium утворюються товстостінні спори. Вони вивільняються у грунт після відмирання кореня та можуть зберігати життєздатність багато років.
Життєвий цикл плазмодіофорних грибів родів Polymyxa і Spongospora подібний до циклу Olpidium.
Гриби передають віруси рослинам двома шляхами. За першого віруси, що перебувають у вільному стані в ґрунті, локалізуються (адсорбуються) на зовнішній оболонці зооспор, які з грунтовими водами пересуваються від однієї рослини до іншої. Джерелом зооспор можуть бути вегетативні спорангії, або спори, що пройшли стадію спокою, а вірусу — інфіковані корені рослин, рослинні рештки. Вважається, що віруси потрапляють у протоплазму зооспори з джгутиком, який утягується під час утворення цисти й на поверхні якого локалізовані віруси. Нагромаджені в такий спосіб у протоплазмі зооспор віруси переносяться в кореневі клітини рослини в процесі переливання вмісту цисти. Зауважимо, що такий шлях інфікування рослин притаманний тільки грибам роду Olpidium. Прикладом інфікування рослин цим способом є передавання вірусу некрозу огірка Olpidium bomovanus, некрозу тютюну Olpidium brassicae.
Інший шлях інфікування рослин грибами — за участю спор. Гриб набуває віруси з рослинної клітини, коли він у ній розвивається. Вони зберігаються в товстостінних спорах. Спори, що перебувають у стані спокою, залишаються життєздатними та зберігають віруси протягом кількох років. Передавання вірусів кореням рослин у цьому разі здійснюється, як і в попередньому, зооспорами. Цей спосіб передавання вірусів грибами рослинам притаманний усім трьом родам — Olpidium, Polymyxa і Spongospora. Прикладами такого шляху інфікування рослин є передавання вірусу призупинення росту тютюну грибами Olpidium brassicae, жовтої мозаїки ячменю грибами Polymyxa graminis, моп-топ вірусу картоплі грибами Spongospora subterranea.
Поміж численних вірусів грибів лише незначна їх кількість зовні подібна до вірусів рослин. У зв’язку з цим виникає припущення про
те, що гриби слугують у більшості випадків інфікування рослин лише місцем утримання (резервуаром) для вірусів рослин. Справді, вірус тютюнової мозаїки був виділений з конідій збудників справжньої борошнистої роси Sphaerotheca lanestris та Erysyphe graminis. Відомі також інші приклади локалізації вірусів рослин у грибах.