ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 1. ОСНОВЫ БИОХИМИИ СТРОЕНИЕ И КАТАЛИЗ - 2011
1. ОСНОВЫ БИОХИМИИ
1.5. Эволюционные основы биохимии
Биологические явления обретают смысл, если их рассматривать через призму эволюции.
Феодосий Добжанский, преподаватель биологии, март 1973
Огромный прогресс в биохимии и молекулярной биологии, произошедший в последние десятилетия, лишь подтвердил замечательное обобщение Добжанского. Удивительное сходство метаболических процессов и последовательностей генов у самых разных организмов строго доказывает, что все существующие сегодня организмы имеют общего эволюционного предшественника и произошли от него в результате ряда небольших изменений (мутаций), каждое из которых обеспечивало определенному организму преимущество в конкретной экологической нише.
Изменения наследственной информации создают возможность для эволюции
Несмотря на практически безукоризненную точность репликации, иногда в этом процессе случаются ошибки, приводящие к изменениям нуклеотидной последовательности ДНК, т. е. генетическим мутациям (рис. 1-32), изменяющим генетическую программу того или иного компонента клетки. Неверно исправленное повреждение одной из цепей ДНК приводит к такому же неправильному результату. Передаваемые по наследству мутации ДНК, т. е. мутации в репродуктивных клетках, могут оказаться для нового организма или клетки опасными, или даже летальными. Например, мутация может привести к синтезу аномального фермента, который не способен катализировать важную метаболическую реакцию. Однако иногда возникают мутации, помогающие организмам или клеткам выживать в определенных условиях. Мутантный фермент, например, может иметь несколько иную специфичность, позволяющую ему использовать субстрат, который клетка ранее не могла метаболизировать. Если популяция мутантных клеток окажется в условиях, когда этот субстрат является единственным или преобладающим, она получит преимущество перед остальными немутировавшими клетками (клетками дикого типа). Мутантная клетка и ее потомство выживут и будут процветать в таких новых условиях, а клетки дикого типа будут голодать и постепенно исчезнут. Этот феномен Дарвин назвал «выживанием наиболее приспособленного организма в условиях отбора», по сути это процесс природной селекции.
Рис. 1-32. Репликация ДНК и мутации: возможный путь появления новой ферментативной активности. В данном примере рассматривается возможность случайного события, когда ген гексокиназы в гипотетическом организме при репликации ДНК копируется дважды. В результате такой организм несет в себе две полные копии гена гексокиназы, одна из которых не нужна. В ходе множества клеточных делений ДНК двух генов гексокиназы продолжает реплицироваться, причем в этом процессе возможны случайные редкие ошибки, приводящие к изменениям последовательности одной копии и кодируемого ею белка. Очень редкое, но возможное событие: измененный белок получает способность связываться с новым субстратом (галактоза в нашем гипотетическом случае). Несущая такой мутантный ген клетка теперь обладает способностью метаболизировать галактозу, что позволит ей выжить в тех условиях, когда глюкозы в среде нет, но есть галактоза. Если бы репликация гена не предшествовала его мутации, была бы утеряна исходная функция данного белка (метаболизм глюкозы).
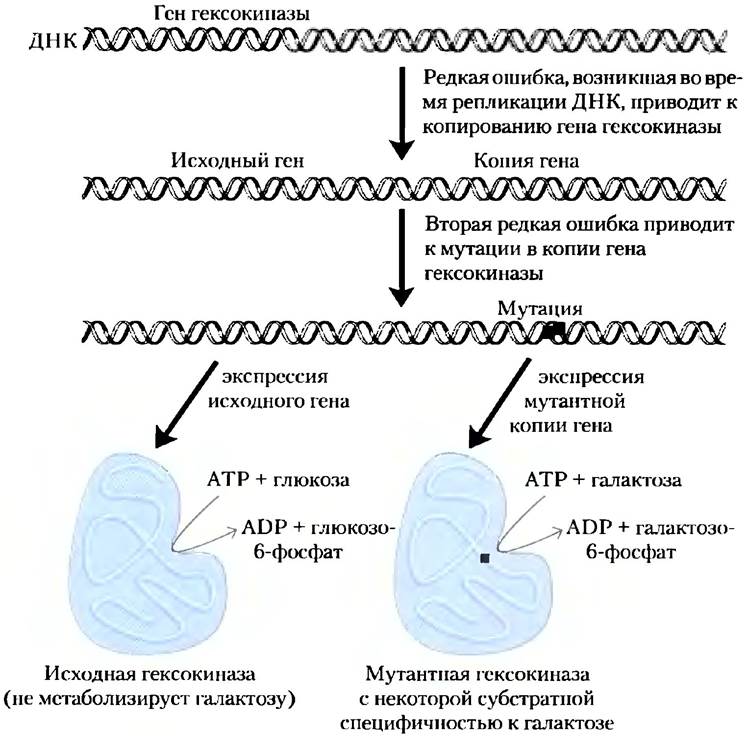
В результате ошибки при репликации хромосомы в нее может встраиваться вторая полная копия гена. Вторая копия является избыточной, и такие мутации не очень опасны. Это один из способов, позволяющий клеткам эволюционировать: возникает новый ген с новыми функциями при сохранении исходного гена с прежними (обычными) функциями. С этой точки зрения молекулы ДНК современных организмов являются историческими документами, запечатлевшими долгий путь от самых первых клеток к сегодняшним видам. Однако исторический «отчет» ДНК неполный: в ходе эволюции многие мутации должны были исчезнуть. Но гем не менее молекулы ДНК являются лучшим документом истории биологии, имеющимся в нашем распоряжении. Частота ошибок при репликации ДНК не может быть слишком высокой, поскольку это привело бы к нежизнеспособности следующего поколения клеток, но не может быть и слишком низкой, поскольку тогда исключается возможность генетической изменчивости, позволяющей выжившим мутантам приспосабливаться к новым условиям обитания.
Несколько миллиардов лет адаптивного отбора научили клетки извлекать максимум выгоды из химических и физических свойств молекул потребляемых веществ. Удачные генетические изменения в отдельных организмах популяции в сочетании с естественным отбором (выживанием и воспроизведением организмов, наилучшим образом приспособившихся к изменившимся условиям обитания) привели к возникновению огромного разнообразия современных организмов, каждый из которых адаптирован к существованию в определенной экологической нише.
Биомолекулы возникли в процессе химической эволюции
Итак, мы познакомились с первой главой истории эволюции — возникновением первых живых клеток. Органические вещества, в том числе основные биомолекулы типа аминокислот и углеводов, из которых состоят живые организмы, обнаруживаются в земной коре, океанах и в атмосфере лишь в следовых количествах. Каким же образом у первых живых организмов возникли столь характерные органические соединения, используемые в качестве строительных блоков?
В соответствии с одной из гипотез эти соединения возникли в результате мощных атмосферных воздействий (ультрафиолетового излучения, вспышек молнии или вулканических извержений) на газы в пребиотической атмосфере Земли и на неорганические вещества в горячих источниках в глубинах океана.
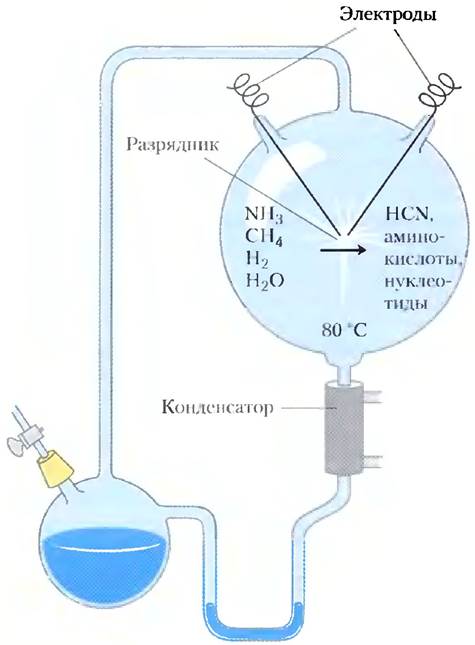
Классический опыт, иллюстрирующий абиотическое (небиологическое) происхождение органических биомолекул, был осуществлен в 1953 г. Стенли Миллером в лаборатории Гарольда Юри. Миллер на протяжении недели или более пропускал электрические разряды, имитировавшие молнии, через газовую смесь, подобную существовавшей на Земле перед зарождением жизни, которая содержала метан, аммиак, водяные пары и водород, заключенную в пространстве между двумя электродами (рис. 1-33), а затем анализировал продукты в закрытом реакционном сосуде. В газовой фазе были обнаружены оксид и диоксид углерода, а также исходные вещества. Водная фаза содержала разнообразные органические соединения, в том числе некоторые аминокислоты, гидроксикислоты, альдегиды и цианистый водород НСN. Этот эксперимент доказал возможность абиотического пути образования биомолекул в относительно мягких условиях за сравнительно короткий срок.
Рис. 1-33. Абиотический синтез биомолекул. Искроразрядный аппарат, использовавшийся Миллером и Юри, в экспериментах по демонстрации абиотического образования органических веществ в условиях примитивной атмосферы. Газовая смесь внутри сосуда подвергалась электрическому воздействию, после чего продукты реакции конденсировались и анализировались. Среди продуктов были обнаружены различные биомолекулы, в том числе и аминокислоты.
Более сложные лабораторные эксперименты предоставили доказательства того, что многие химические составляющие живых клеток, в том числе полипептиды и РНК-подобные молекулы, также могут образовываться в этих условиях. Полимеры РНК могут проявлять каталитические свойства в важных биологических реакциях (с.м. гл. 26 и 27). Кроме того, РНК, вероятно, играла важнейшую роль в пребиотической эволюции как в качестве катализатора, так и в качестве источника информации.
РНК и схожие с ней предшественники могли быть первыми генами и катализаторами
В существующих сегодня организмах нуклеиновые кислоты кодируют генетическую информацию, определяющую структуру ферментов, а ферменты катализируют репликацию и репарацию нуклеиновых кислот. Взаимозависимость этих двух классов соединений ставит трудный вопрос: что было раньше — ДНК или белок?
Однако может быть, что они появились одновременно и им обоим предшествовала РНК. Открытие того факта, что молекулы РНК способны катализировать собственный синтез, указывает на возможную роль РНК или подобных молекул в качестве первых генов и первых катализаторов. В соответствии с данным сценарием (рис. 1-34) на одной из ранних стадий биологической эволюции в «первичном бульоне» возникла молекула РНК, которая смогла катализировать синтез других молекул РНК с той же последовательностью, т. е. образовалась самореплицирующаяся молекула РНК. Концентрация молекул РНК должна была возрастать экспоненциально, поскольку из одной молекулы получается две, из двух — четыре и т. д. Точность саморепликации, вероятно, была не абсолютной, поэтому стали появляться варианты РНК, некоторые из которых оказать еще лучше приспособлены к саморепликации. В конкурентной борьбе за нуклеотиды победили наиболее эффективные молекулы, а менее эффективные исчезли из популяции.
Рис. 1-34. Возможный сценарий развития «мира РНК».
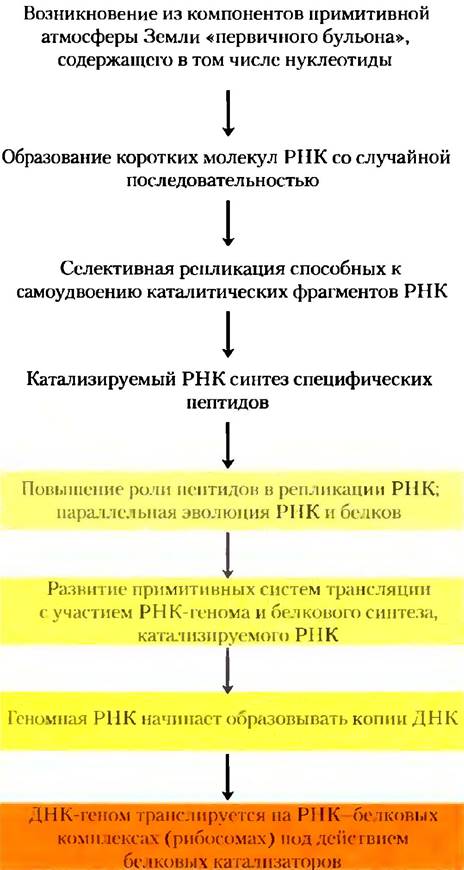
В соответствии с гипотезой «мира РНК» разделение функций между ДНК (хранение генетической информации) и белком (катализ) произошло позднее. Возникали новые варианты самореплицирующихся РНК, которые были способны также катализировать конденсацию аминокислотных звеньев в полипептидные цепи. В какой-то момент образовавшиеся таким образом пептиды стали усиливать способность РНК к саморепликации. Такая пара РНК/пептид- помощник могла претерпевать дальнейшие изменения, создавая еще более эффективные системы репликации. Замечательное открытие того факта, что в аппарате для синтеза белка современной клетки (рибосоме) не белок, а РНК катализирует образование пептидной связи, свидетельствует в пользу гипотезы «мира РНК».
Через какое-то время после возникновения этих примитивных систем для белкового синтеза произошло следующее изменение: функцию сохранения «генетической» информации взяла на себя молекула ДНК, последовательность которой комплементарна последовательности самореплицирующейся РНК. Молекулы РНК, в свою очередь, эволюционировали, совершенствуя катализ синтеза белка. (Далее в гл. 8 мы остановимся на том, почему молекула ДНК более стабильна по сравнению с РНК и лучше подходит для хранения наследственной информации.) Белки проявили себя в качестве универсальных катализаторов и позднее взяли на себя эту функцию. Липидоподобные компоненты из «первичного бульона» образовали относительно непроницаемые слои, окружавшие самореплицирующиеся ассоциации молекул. Высокая концентрация белков и нуклеиновых кислот внутри этих липидных оболочек способствовала молекулярным взаимодействиям, необходимым для репликации.
Биологическая эволюция началась более трех с половиной миллиардов лет назад
Земля возникла около 4,6 млрд лет назад, а в соответствии с имеющимися данными первые живые организмы появились более 3,5 млрд лет назад. В 1996 г. работавшие в Гренландии ученые обнаружили не окаменевшие останки, а химические доказательства жизни («топливные молекулы»), возраст которых свыше 3,85 млрд лет. Они нашли вросшие в камень углеродсодержащие вещества, вероятно, биологического происхождения. В первый миллиард лет существования Земли кое-где стали появляться первые простые организмы, способные воспроизводить свою собственную структуру на основании матрицы (РНК?), служившей первым генетическим материалом. Поскольку атмосфера Земли в те времена была практически лишена кислорода, и лишь немногие микроорганизмы использовали органическое вещество, образовавшееся в естественных процессах, существовавшие органические соединения были относительно устойчивыми. С учетом этой устойчивости органических соединений и огромной длительности происходивших процессов, можно понять, каким образом невероятное стало неизбежным: органические вещества включались в развивающиеся клетки, эффективность механизмов самовоспроизведения возрастала. Начался процесс биологической эволюции.
Первые клетки, вероятно, были хемогетеротрофами
Самые первые клетки возникали в восстановительной атмосфере (где нет кислорода) и, вероятно, получали энергию из неорганических веществ, таких как сульфид железа (II) и карбонат железа (II), которые тогда на Земле имелись в большом количестве. Например, в результате реакции
FeS + H2S —> FeS2 + Н2
выделяется достаточное количество энергии, чтобы обеспечить синтез молекулы АТР или аналогичного вещества. Необходимые клеткам органические вещества могли появиться из компонентов ранней атмосферы Земли (СО, СО2, N2, NH3, СН4 и др.) в результате небиологических воздействий, таких как вспышки молнии и вулканические извержения, или в результате гидротермальных процессов. Существует также гипотеза, что органические вещества были привнесены на Землю из космоса. В 2006 г. космический зонд Stardust вернулся на Землю с частицами космической пыли, взятой из хвоста кометы; в этой пыли содержались различные органические вещества.
Первые одноклеточные организмы, возникшие в богатой смеси органических веществ («первичном бульоне»), практически наверняка были хемогетеротрофами (рис. 1-5). Необходимые им органические вещества исходно возникли из компонентов первичной атмосферы (СО, СО2, N2, СН4 и др.) не биологическим путем, а под действием тепла от вулканической деятельности и разрядов молний. Первые гетеротрофы постепенно приобретали способность добывать энергию из окружающих веществ, использовали ее для синтеза необходимых им молекул и становились менее зависимыми от внешних источников питания. Очень важным шагом в эволюции было появление пигментов, способных захватывать энергию Солнца, с помощью которой клетки могли восстанавливать («фиксировать») СО2, превращая его в более сложные органические вещества. Первым донором электронов для этого процесса фотосинтеза, по-видимому, был H2S, превращавшийся в элементарную серу или сульфат-ионы (SO2-4). Позднее клетки стали использовать в качестве донора электронов воду, в результате чего в атмосферу начал выделяться кислород. Потомками тех первых фотосинтезирующих организмов, продуцирующих кислород, являются современные цианобактерии.
Поскольку атмосфера в ранние периоды существования Земли почти не содержала кислорода, первые клетки были анаэробами. В таких условиях хемотрофы могли окислять органические вещества до СО2, передавая электроны не па кислород, а на такие акцепторы, как SО2-4, в результате чего в качестве побочного продукта образовывался H2S. С появлением продуцирующих кислород фотосинтезирующих бактерий атмосфера все более обогащалась кислородом — мощным окислителем и смертельным ядом для анаэробов. В этих сложных условиях, которые Линн Маргелис и Дорион Саган назвали «кислородным холокостом», некоторые линии микроорганизмов дали начало аэробам, получавшим энергию в процессе передачи электронов от топливных молекул кислороду. Поскольку такие реакции сопровождаются высвобождением значительного количества энергии, в аэробных условиях использующие кислород организмы получили энергетическое преимущество над анаэробными собратьями. В результате в кислородсодержащей атмосфере стали преобладать аэробные организмы.
Современные бактерии населяют практически все экологические пиши в биосфере; существуют организмы, способные использовать в качестве источника углерода и энергии практически любой тип органических веществ. Фотосинтезирующие микробы в пресной и в соленой воде захватывают солнечную энергию и используют ее для образования углеводов и других клеточных компонентов, которые, в свою очередь, служат источником питания для других живых организмов. Процесс эволюции продолжается. С помощью быстро воспроизводящихся бактериальных клеток мы можем быть свидетелями этого процесса в лабораторных условиях. Один из подходов к получению примитивной клетки («протоклетки») в лабораторных условиях состоит в том, чтобы определить минимальное количество необходимых для жизни генов путем анализа геномов самых просто устроенных бактерий. Минимальным геномом среди свободноживущих бактерий обладает Mycobacterium genitalium: ее геном состоит из 580 000 пар нуклеотидов и содержит 483 гена.
Эукариотические клетки возникли в несколько стадий из более простых предшественников
Судя по ископаемым остаткам, около 1,5 млрд лет назад начали возникать более крупные и сложные организмы, возможно, это были первые эукариотические клетки (рис. 1-35). Детали эволюционного пути от доядерных к ядерным клеткам невозможно установить только на основании ископаемых остатков, однако морфологическое и биохимическое изучение современных организмов позволило приблизительно представить последовательность событий, согласующихся с результатами анализа ископаемых остатков.
Рис. 1-35. Этапы эволюции жизни на Земле.

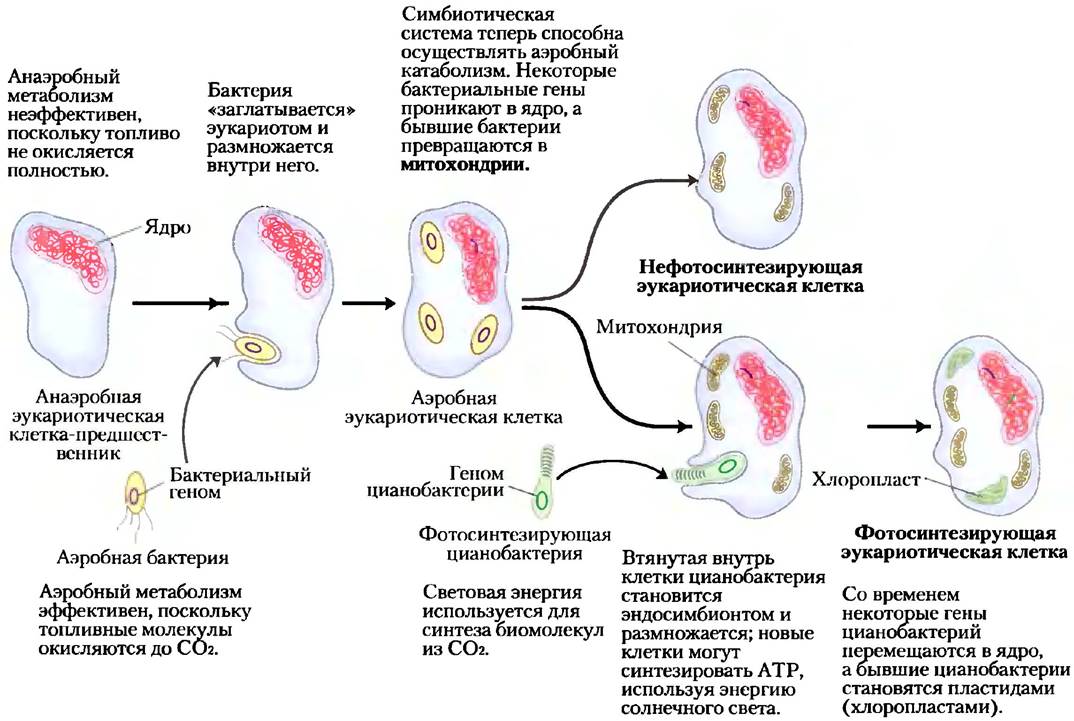
Обязательно должны были произойти три принципиальных изменения. Во-первых, поскольку клетка обзаводилась все большим количеством ДНК, нужны были более сложные механизмы для осуществления компактной упаковки ДНК в комплексах со специфическими белками, а также для точного разделения между дочерними клетками в процессе деления. Для этого понадобились специализированные белки, стабилизирующие свернутую ДНК и разрушающие комплексы ДНК с белком (хромосомы) при делении клетки. Во-вторых, при увеличении размера клетки стала развиваться система внутренних мембран, в том числе появилась двойная мембрана, окружающая ДНК. Эта мембрана отделяла синтез РНК на основе ДНК-матрицы, протекающий в ядре, от синтеза белка на рибосомах в цитоплазме. Наконец, в-третьих, ранние эукариотические клетки, которые не могли осуществлять фотосинтез или аэробный метаболизм, включали в себя аэробные или фотосинтезирующие бактерии, образуя эндосимбиотические ассоциации, со временем ставшие постоянными (рис. 1-36). Некоторые аэробные бактерии превратились в митохондрии современных эукариот, а некоторые фотосинтезирующие цианобактерии стали пластидами, как, например, хлоропласты в зеленых водорослях — вероятных предшественниках клеток современных растений.
Рис. 1-36. Роль эндосимбиоза в эволюции эукариот. Самые первые эукариоты были анаэробами. Они включили внутрь себя пурпурные бактерии (показаны желтым цветом), которые придали им способность осуществлять аэробный катаболизм и со временем превратились в митохондрии. Фотосинтезирующие цианобактерии (показаны зеленым цветом) впоследствии стали эндосимбионтами некоторых аэробных эукариот, превратив эти клетки в фотосинтезирующих предшественников современных зеленых водорослей и растений.
На более поздних этапах эволюции одноклеточные организмы стали образовывать кластеры, что позволило им увеличить подвижность, повысить эффективность метаболизма или репродукции по сравнению со свободноживущими одиночными клетками. Дальнейшая эволюция таких кластеров привела к возникновению устойчивых ассоциаций отдельных клеток и постепенному развитию специализации внутри колоний - произошла дифференцировка клеток.
Использование преимуществ клеточной специализации привело к возникновению еще более крупных и высокодифференцированных организмов, в которых одни клетки осуществляли сенсорные функции, другие — пищеварительные, фотосинтетические, репродуктивные и т. д. Существующие сейчас многоклеточные организмы содержат сотни клеток различного типа, каждый из которых специализируется на определенной функции, обслуживающей весь организм. Основные механизмы, возникшие на заре эволюции, развивались и оттачивались далее. В основе пульсирующих движений ресничек парамеции и жгутиков хламидомонады лежат те же самые ключевые структуры и механизмы, что используются, например, высокоспециализированными клетками сперматозоидов позвоночных.
Молекулярное строение раскрывает эволюционные связи
Сегодня биохимики обладают огромной и все пополняющейся сокровищницей знаний о молекулярном строении клеток, которые можно использовать для анализа эволюционных связей и усовершенствования эволюционной теории; и количество знаний все увеличивается. Последовательность генома (всего генетического багажа организма) полностью определена уже для сотен бактерий, более 40 архей, постоянно увеличивающегося числа эукариотических микроорганизмов (например, Saccharomyces cerevisiae и Plasmodium sp.), растений, включая Arabidopsis thaliana и рис, многоклеточных животных, включая Сае- norhabditis elegans (круглый червь), Drosophila melanogaster(фруктовая мушка), мышь, крысу, собаку, шимпанзе и Homo sapiens (табл. 1-2). Этот список постоянно пополняется. Зная последовательности геномов, можно проводить детальное сравнение различных видов и глубже понять эволюционный процесс. До настоящего времени молекулярная систематика, основанная на последовательностях генов, согласовывалась с классической систематикой, основанной на изучении макроскопических структур, но во многом была более точна. Хотя на уровне анатомии постоянно происходило расхождение организмов, основополагающее единство живого становится очевидным при проведении исследований на молекулярном уровне. Молекулярные структуры и механизмы самых простых и самых сложных организмов обладают замечательным сходством. Эти сходства легче всего проследить на уровне последовательностей либо ДНК, кодирующих белки, либо самих белков.
Таблица 1-2. Некоторые организмы, для которых определены нуклеотидные последовательности
Организм |
Размер генома (млн пар нуклеотидов) |
Количество генов |
Биологические особенности |
Mycoplasma genitaliim |
5,8 х 105 |
4,8 x 102 |
Вызывает пневмонию |
Treponema pallidum |
1,1 х 106 |
1,0 x103 |
Вызывает сифилис |
Borrelia burgdorferi |
9,1 х 105 |
8,5 x 102 |
Вызывает болезнь Лайма |
Helicobacter pylori |
1,7 x 106 |
1,6 x 103 |
Вызывает язву желудка |
Methanococcus jannaschii |
1,7 x 106 |
1,7 x 103 |
Растет при 85 °С! |
Haemophilus influenzae |
1,8 x 106 |
1,6 x 103 |
Вызывает грипп |
Archaeoglobus fulgidus |
2,2 x 106 |
2,4 x 103 |
Относится к царству архей, метаноген |
Synechocystis sp. |
3,6 x 106 |
3,2 x 103 |
Цианобактерия |
Bacillus subtilis |
4,2 x 106 |
4,1 x 103 |
Распространенная почвенная бактерия |
Escherichia coli |
4,6 x 106 |
4,4 x 103 |
Некоторые штаммы вызывают токсический шок |
Saccharomyces cerevisiae |
1,2 x 107 |
5,9 x 103 |
Одноклеточный эукариотический организм |
Plasmodium falciparum |
2,3 x 107 |
5,3 x 103 |
Вызывает малярию у человека |
Caenorhabditis elegans |
1,0 x 108 |
2,3 x 104 |
Многоклеточный круглый червь |
Anopheles gambiae |
2,3 x 108 |
1,3 x 104 |
Переносчик малярии |
Arabidopsis thaliana |
1,2 x 108 |
3,2 x 104 |
Сорное растение |
Oryza sativa |
3,9 x 108 |
3,8 x 104 |
Посевной рис |
Drosophila melanogaster |
1,2 x 108 |
2,0 x 104 |
Фруктовая мушка |
Mus musculus domesticus |
2,6 x 1O9 |
2,7 x 104 |
Лабораторная мышь |
Pan troglodytes |
3,1 x 109 |
4,9 x 104 |
Шимпанзе |
Homo sapiens |
3,1 x 109 |
2,9 x 104 |
Человек |
Источник: страница RefSeq для каждого организма на www.nebi.nlm.nih.gov/genomcs
Если последовательности двух генов (нуклеотидые последовательности ДНК или последовательности кодируемых ими белков) имеют очевидное сходство, то их называют гомологичными, а кодируемые ими белки — гомологами. Если два гомологичных гена обнаружены у одного и того же вида, их называют паралогичными, а кодируемые ими белковые продукты — паралогами. Считается, что паралогичные гены образовались путем удвоения гена и постепенных изменений последовательностей обеих копий. Обычно паралогичные белки имеют не только схожие аминокислотные последовательности, но и аналогичную трехмерную структуру, хотя в ходе эволюции стали выполнять различные функции.
Два гомологичных гена (или белка) из организмов разных видов называют ортологичными, а их белковые продукты — ортологами. Ортологи в обоих организмах обычно выполняют одну и ту же функцию, поэтому, когда выясняется, что вновь секвенированный ген в организме одного вида ортологичен гену из организма другого вида, можно предположить, что они кодируют белки с одинаковыми функциями. Так, без проведения биохимических исследований на основании геномных последовательностей можно определить функции белковых продуктов. Аннотированный геном кроме самой последовательности ДНК содержит описание предполагаемой функции продукта каждого гена, выведенной на основании сравнения данной геномной последовательности с последовательностями из других организмов, в которых функции белков известны. В принципе, определяя метаболические пути (наборы ферментов), кодируемые данным геномом, можно вывести метаболические характеристики организма на основании одной только геномной последовательности.
Различия в последовательностях гомологичных генов можно рассматривать как грубый способ оценки степени расхождения двух видов в процессе эволюции, т. е. эти различия позволяют оценить, как давно общий предшественник дал начало двум эволюционным линиям. Чем больше различий, тем раньше произошло расхождение. Можно построить филогенетическое (фамильное) древо, на котором эволюционная дистанция между двумя любыми видами определяется близостью их расположения (например, рис. 1-4).
В ходе эволюции появляются новые структуры, процессы и регуляторные механизмы, что отражает изменения в геномах эволюционирующих организмов. В геноме такого простого эукариотического организма, каким являются дрожжи, должны были появиться отсутствующие у бактерий и архей гены, отвечающие за образование ядерной мембраны. Геном насекомых должен содержать отсутствующие у дрожжей гены белков, определяющие специфическую сегментированную структуру тела насекомых. Геномы всех позвоночных должны содержать гены, несущие информацию о развитии позвоночника, а геномы млекопитающих — уникальные гены, отвечающие за образование плаценты, существующей только у млекопитающих, и т. д. Сравнение целых геномов организмов каждого типа приводит к выявлению генов, имеющих принципиальное значение с точки зрения фундаментальных эволюционных изменений в структуре и развитии организма.
Функциональная геномика указывает назначение генов в специфических клеточных процессах
После определения полной нуклеотидной последовательности генома и аннотирования каждого гена (т. е. приписывания каждому гену определенной функции) молекулярный генетик может сгруппировать гены в соответствии с теми процессами, в которых они участвуют (синтез ДНК или белка, образование АТР и др.). Таким образом удается определить, какая часть генома отвечает за ту или иную активность клетки. Функции многих генов (более 40%) в клетках Е. coli, A. thaliаnа и Н. sapiens до сих пор неизвестны. У всех трех видов значительную часть генома составляют гены транспортных белков, отвечающих за перемещение ионов и небольших молекул через плазматическую мембрану (их все же больше у бактерий и растений, чем у млекопитающих: 10% из ~4 400 генов у Е. coli, -8% из -32 000 генов у A. thaliana и -4% из -29 000 генов у Н. sapiens). Гены, кодирующие белки и РНК для их синтеза, составляют 3-4% генома Е. coli. В более сложно устроенных клетках А. thaliana больше генов требуется для доставки белков к местам их основной локализации, чем для синтеза самих этих белков (соответственно 6 и 2% генома). В общем, чем сложнее организм, тем большую часть его генома составляют гены, регулирующие клеточные процессы, и меньшая часть отвечает за реализацию основных процессов тина образования АТР и синтеза белка.
Сравнительный анализ геномов играет все большую роль в биологии человека и медицине
Геномы человека и шимпанзе совпадают на 99,9%, но разница между двумя видами огромна. Эти небольшие различия в геномах должны объяснить человеческую способность говорить, чрезвычайную развитость мышц шимпанзе и массу других вещей. Сравнение геномов позволит исследователям идентифицировать гены, которые могут быть связаны с расхождением в программах развития человека и других приматов и с появлением у человека более сложных функций, в том числе языка общения. Картина станет проясняться только по мере того, как будет проведено сравнение генома человека с геномными последовательностями большего числа приматов.
Аналогичным образом, генетические различия между людьми чрезвычайно ничтожны по сравнению с различиями между человеком и шимпанзе, тем не менее все мы очень разные, в том числе с точки зрения здоровья и предрасположенности к хроническим заболеваниям. Еще очень многое предстоит узнать о различиях в геномных последовательностях людей, но со всей определенностью можно предсказать, что в ближайшие десятилетия доступность генетической информации принципиально изменит систему медицинской диагностики и лечения. Можно ожидать, что паллиативное лечение некоторых генетических заболеваний сменится реальной помощью, а болезни, предрасположенность к которым связана с наличием определенных генетических маркеров, можно будет предупреждать усиленными превентивными мерами. Сегодняшняя «история болезни», возможно, сменится медицинским «прогнозом». ■
Краткое содержание раздела 1.5 Эволюционные основы биохимии
■ Наследуемые случайные мутации приводили к появлению организмов, лучше приспособленных к выживанию в определенных экологических нишах, и такое потомство выдержало естественный отбор. Этот процесс мутаций и отбора лежит в основе дарвиновской теории эволюции, указавшей на происхождение всех живущих ныне организмов от первой клетки и объяснившей фундаментальное сходство всех живых организмов.
■ Жизнь возникла около 3,5 млрд лет назад, наиболее вероятно, с появлением заключенной в мембрану самореплицирующейся молекулы РНК. Составляющие первой клетки могли возникнуть вблизи горячих источников в глубинах океана или в результате действия молнии или высокой температуры на такие простые молекулы, как СO2 и NН3.
■ Каталитическая и генетическая функции РНК первых геномов со временем были переданы белкам и ДНК.
■ В результате эндосимбиоза с бактериями эукариотические клетки приобрели способность к фотосинтезу и окислительному фосфорилированию. Возникли многоклеточные организмы с дифференцировкой клеток, в результате которой различные типы клеток специализируются на выполнении одной или нескольких функций, важных для жизнедеятельности всего организма.
■ Знание полных нуклеотидных последовательностей организмов, находящихся на разных ветвях филогенетического древа, способствует пониманию эволюционного процесса и открывает широчайшие возможности для развития медицины.