ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
19. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ И ФОТОФОСФОРИЛИРОВАНИЕ
19.7. Поглощение света
Видимый свет — это электромагнитное излучение с длинами волн от 400 до 700 нм, т. е. небольшая часть спектра солнечного излучения в диапазоне от фиолетового до красного (рис. 19-46). Энергия фотонов (квантов световой энергии) обратно пропорциональна длине световой волны. Фотоны фиолетовой (коротковолновой, более высокочастотной) области видимого спектра несут более высокую энергию по сравнению с фотонами красной области. Энергия (Е) одного фотона в видимой области спектра можно рассчитать по уравнению Планка:
Е = hv = hc/λ
где h — постоянная Планка, h = 6,625 • 10-34 Дж • с, v — волновое число, частота в с-1, с — скорость света, с = 3,00 • 108 м/с, λ — длина волны (в метрах). Энергия фотона видимого света различна в зависимости от его длины волны (частоты): в красной области -150 кДж/эйнштейн, в фиолетовой области -300 кДж/эйнштейн.
Рис. 19-46. Электромагнитное излучение. Спектр электромагнитного излучения и энергия фотонов в видимой области спектра. 1 Эйнштейн = 6 • 1023 фотонов.
Способность химического соединения поглощать свет зависит от распределения электронов вокруг ядер в его молекуле. При поглощении молекулой (хромофором) одного фотона один из ее электронов переходит на более высокий энергетический уровень. Происходит это по закону «все или ничего»: чтобы перевести электрон на более высокий энергетический уровень, фотон должен обладать определенным количеством энергии (квантом, от лат. quantum — количество), точно соответствующем энергии перехода электрона. Молекула, поглотившая один фотон, находится в высокоэнергетическом, возбужденном состоянии, которое, как правило, неустойчиво. Если отключить источник света, электроны с высокоэнергетических орбиталей обычно быстро вновь переходят на свои низкоэнергетические уровни; при этом молекула возвращается в исходное стабильное, так называемое основное состояние, высвобождая энергию возбуждения (в форме света или тепла). Флуоресценция — свет, испускаемый возбужденной молекулой при ее возвращении в основное состояние; длина волны испускаемого при флуоресценции света всегда больше длины волны поглощенного света (см. доп. 12-3, т. 1). Другой тип перехода возбужденной молекулы в исходное состояние, очень важный для фотосинтеза, заключается в прямой передаче энергии возбуждения от возбужденной молекулы к соседней, находящейся в основном состоянии. Такой процесс передачи энергии называется передачей экситона, а квант энергии, переданной от одной молекулы к другой, называется экситоном, по аналогии с фотоном — квантом энергии света.
Пример 19-2 Энергия фотона
Сосудистые растения при фотосинтезе используют свет с длиной волны около 700 нм. Найдите энергию «моля» фотонов (эйнштейна фотонов) света с этой длиной волны и сравните полученную величину с энергией, необходимой для синтеза 1 моль АТР.
Решение. Энергия одного фотона определяется по уравнению Планка. При длине волны 700 • 10-9 м энергия одного фотона равна
Е = hc /λ = [(6,626 • 10-34 Дж • с)(3,00 • 108 м/с)]/(7,00 • 10-7 м) = 2,84 •10-19 Дж
1 эйнштейн = 1 моль фотонов, что соответствует числу фотонов, равноиу числу Авогадро (6,022 • 1023). Отсюда можно рассчитать энергию одного эйнштейна фотонов при 700 нм:
(2,84 • 10-19 Дж/фотон) (6,022 • 1023 фотон/эйнштейн) = 17,1 • 104 Дж/эйнштейн = 171 кДж/эйнштейн
Таким образом, 1 моль фотонов красного света обладает энергией, которая примерно в 5 раз превышает энергию образования 1 моль АТР из ADP и Pi (30,5 кДж/моль).
При фотосинтезе энергию света поглощают хлорофиллы
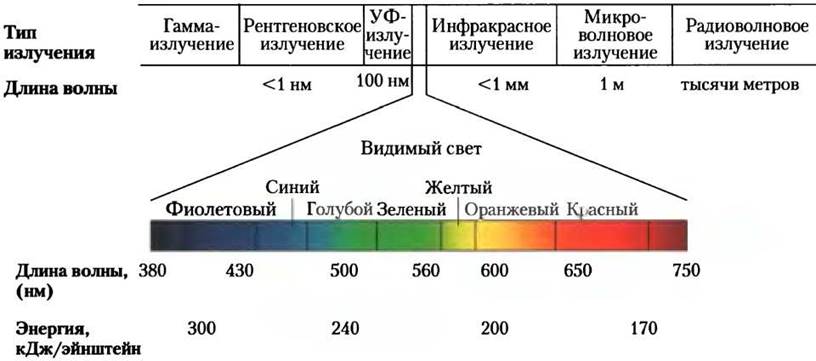
Обратимся теперь к рассмотрению светопоглощающих пигментов тилакоидных мембран. Главную роль среди них играют зеленые пигменты хлорофиллы —Mg2+-содержащие молекулярные комплексы с полициклической плоской структурой, как у протопорфирина гемоглобина, но комплексообразователем является не Fe2+, а Mg2+ (см. рис. 5-1, т. 1). Четыре атома азота, обращенные в центр кольцевой структуры, координационно связаны с ионом Mg2+ (рис. 19-47). У всех хлорофиллов длинная изопреноидная боковая цепь — это остаток спирта фитола, присоединенного сложноэфирной связью к карбоксильной группе заместителя в кольце IV. В хлорофиллах, в отличие от гемов, есть пятое (непиррольное)кольцо.
Пятичленные гетероциклы вокруг иона Mg2+ образуют сопряженную систему двойных и одинарных связей, которая дает интенсивные характеристические полосы в видимой области спектра, что очень важно для фотосинтеза (рис. 19-48). У хлорофиллов необычно высокие коэффициенты экстинкции (см. доп. 3-1, т. 1), а значит во время фотосинтеза они хорошо поглощают в видимой области спектра.
В хлоропластах всегда присутствуют хлорофиллы двух типов — а и b (рис. 19-47, а). Хотя оба они окрашены в зеленый цвет, их спектры поглощения слегка различаются (рис. 19-48). У большинства высших растений количество хлорофилла а примерно вдвое превышает количество хлорофилла b. Хлорофиллы водорослей и фотосинтезирующих бактерий слегка отличаются от хлорофиллов высших растений.
Молекулы хлорофилла всегда связаны со специальными белками, образуя светопоглощающие комплексы, в которых они определенным образом ориентированы относительно друг друга и к остальным белковым комплексам в мембране тилакоидов. Один светопоглощающий комплекс (тип II; рис. 19-49) содержит семь молекул хлорофилла а, пять молекул хлорофилла b и две молекулы вспомогательного пигмента лютеина (см. ниже).
У цианобактерий и красных водорослей рецепторы световой энергии представлены пигментами другого класса — это красные и синие фикобилины (фикоэритробилины и фикоцианобилины соответственно; рис. 19-47, б). Фикобилины — тетрапирролы с открытой цепью и с большим числом сопряженных двойных связей. В отличие от хлорофилла фикобилины не содержат Mg2+ и не имеют циклической тетрапиррольной структуры. Фикобилипротеинами называются комплексы фикобилинов со специальными белками. Они организованы в крупные комплексы — фикобилисомы (рис. 19-50), которые служат главными светопоглощающими системами у этих микроорганизмов.
Вспомогательные пигменты расширяют диапазон длин волн поглощаемого света
В мембранах тилакоидов присутствуют наряду с хлорофиллами также второстепенные, или вспомогательные, светопоглощающие пигменты. К вспомогательным пигментам относятся различные каротиноиды, окрашенные в желтый, красный и пурпурный цвета. Среди них наиболее важны красный пигмент изопреноидного строения β-каротин и желтый каротиноид лютеин (рис. 19-47, в и г). Каротиноиды поглощают свет в другом диапазоне длин волн, чем хлорофиллы, и поэтому функционируют как световые рецепторы, дополняющие хлорофиллы (рис. 19-48).
Рис. 19-47. Главные и вспомогательные фотопигменты, а — хлорофиллы а и b и бактериохлорофилл — главные светопоглощающие пигменты, б — фикоэритробилин и фикоцианобилин (группа фикобилинов) — антенные, или светособирающие, пигменты цианобактерий и красных водорослей, в — β-каротин и другие каротиноиды, а также лютеин (ксантофилл) — вспомогательные пигменты зеленых листьев. Все эти пигменты содержат систему сопряженных связей, которая дает им способность поглощать свет в видимой области (выделена розовым цветом).
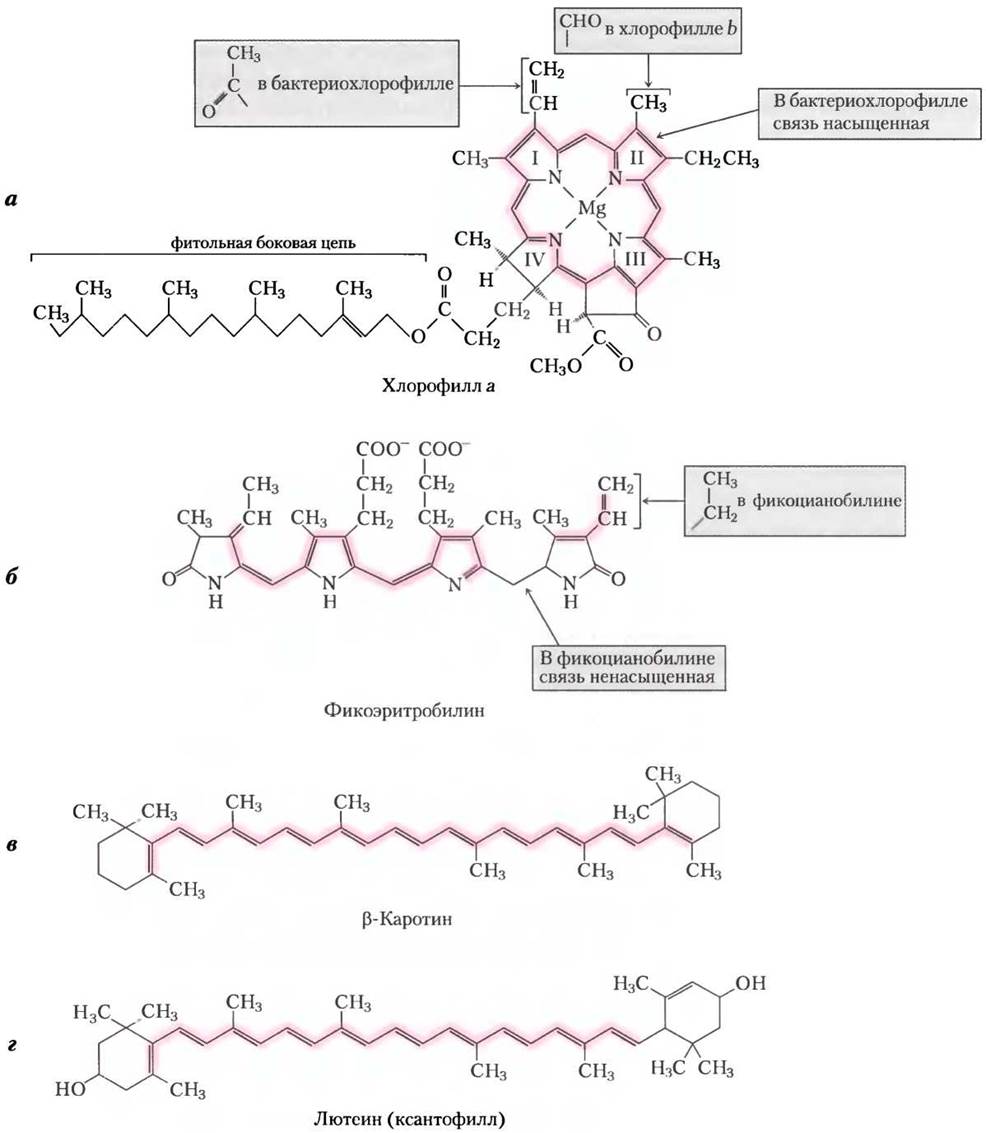
Экспериментальные данные по оценке эффективности фотосинтеза в зависимости от длины волны представляют в виде фотохимического спектра действия (рис. 19-51), который может быть использован также для идентификации главных пигментов, ответственных за биологическую активность света определенной длины волны. Благодаря способности поглощать световую энергию в определенной области длин волн фотосинтезирующие организмы могут занимать свою уникальную экологическую нишу. Например, пигменты фикобилины красных водорослей и цианобактерий поглощают свет в диапазоне 520-630 нм (рис. 19-48), что позволяет им выживать в окружении организмов, пигменты которых и сама вода поглощают свет в более коротковолновом или более длинноволновом диапазоне.
Рис. 19-48. Спектры поглощения фотопигментов в видимой области. Растения имеют зеленую окраску из-за того, что их пигменты поглощают видимый свет в красной и голубой областях, а зеленый свет отражают или пропускают. Сравнение спектра солнечного излучения, достигающего земной поверхности, со спектрами поглощения растительных пигментов показывает, что хлорофиллы а и b и вспомогательные пигменты позволяют растениям поглощать основные количества поступающей на Землю солнечной энергии. Соотношение между хлорофиллами и различными каротиноидами у разных видов растений заметно варьирует, что и определяет характерную зеленую окраску фотосинтезирующих клеток, которая изменяется от сине-зеленой, как у хвоинок ели, или ярко-зеленой, как у листьев клена, до красной, бурой и даже пурпурной, как у разных видов многоклеточных водорослей и листьев некоторых декоративных растений.
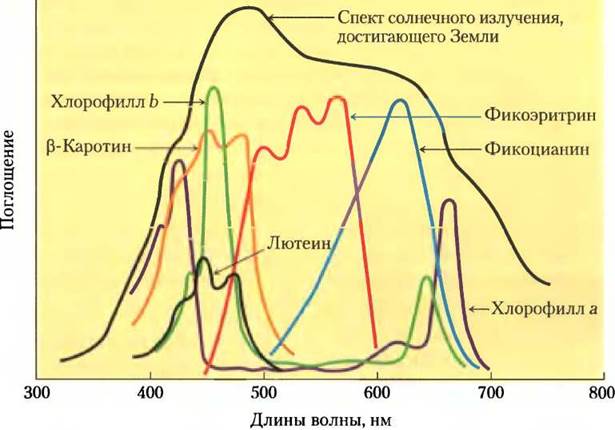
Рис. 19-49. Строение светопоглощающего комплекса типа II (РDВ ID 2ВHW). Функциональный светопоглощающий комплекс представляет собой тример, содержащий 36 молекул хлорофилла и 6 молекул лютеина. В состав представленного здесь мономера (вид в плоскости мембраны) входят три сегмента α-спирали, пронизывающих мембрану насквозь, семь молекул хлорофилла а (светло- зеленые), пять молекул хлорофилла b (темно-зеленые) и две молекулы вспомогательного пигмента лютеина (желтые), образующих внутренний каркас.
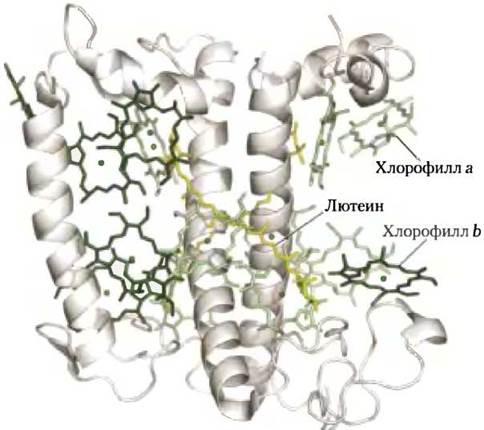
Рис. 19-50. Строение фикобилисомы. Фикобилисома цианобактерий и красных водорослей включает комплексы фикобилинов со специальными белками — фикоэритрином (ФЭ), фикоцианином (ФЦ) и аллофикоцианином (АФ). Энергия фотонов, поглощенная фикоэритрином и фикоцианином, быстро передается в виде экситона через белковый комплекс фикоцианобилина аллофикоцианинхлорофилл а (в реакционном центре). Подробнее этот вопрос обсуждается в тексте.
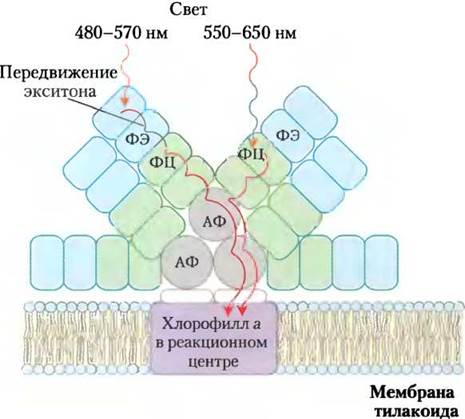
Рис. 19-51. Два метода изучения фотохимического спектра действия, а — результаты классического эксперимента, проведенного в 1882 г. Т. В. Энглеманном, по определению длины волны, наиболее эффективной для фотосинтеза. Волокна фотосинтезирующей морской водоросли поместили на предметное стекло микроскопа и освещали светом, прошедшим через диспергирующую призму, причем на одну часть волокон попадала синяя область спектра, на другую часть — желтая, на третью, — красная. На это же стекло помещали бактерии, обладающие свойством мигрировать к с наиболее высокой концентрации кислорода. Эффективность видимого света данной длины волны оценивали числом скопившихся на участке бактерий — лучшие результаты обеспечивали фиолетовый и красный свет. б — результаты аналогичного эксперимента, отличающегося от предыдущего тем, что количество выделившегося при фотосинтезе кислорода оценивали с помощью кислородного электрода. Фотохимический спектр действия показывает зависимость относительной эффективности фотосинтеза от длины волны. Сравнение спектров поглощения пигментов и фотохимического спектра действия фотосинтезирующих клеток позволяет оценить светопоглощающие свойства пигментов (см. рис. 19-48).

Хлорофиллы передают энергию света реакционным центрам путем переноса экситона на соседние молекулы
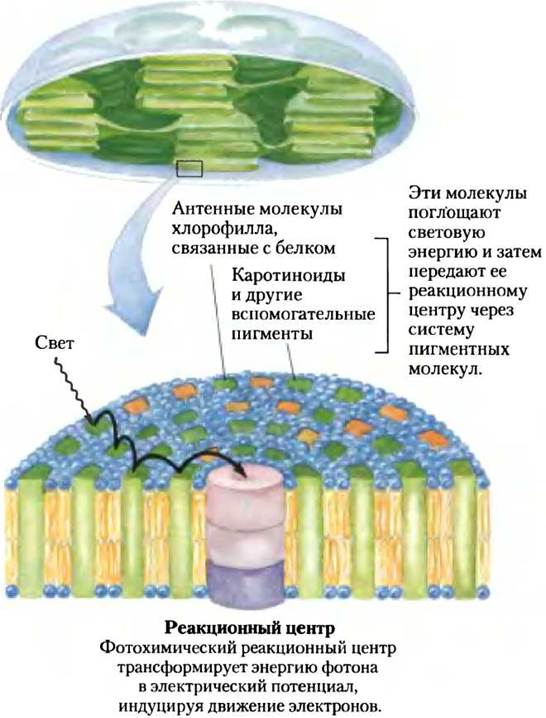
Светопоглощающие пигменты тилакоидных мембран собраны в функциональные комплексы фотосистемы. Фотосистема, например, в хлоропластах шпината содержит около 200 молекул хлорофиллов и около 50 молекул каротиноидов. Молекулы всех пигментов поглощают фотоны, но только некоторые молекулы хлорофиллов могут превращать свет в химическую энергию. Эти специализированные хлорофиллы связаны с особыми белками и образуют фотохимический реакционный центр. Все прочие молекулы пигментов называются светособирающими или антенными. Они поглощают свет и очень быстро передают его энергию фотохимическим реакционным центрам, где и происходит фотохимическая реакция (рис. 19-52).
Рис. 19-52. Строения фотосистемы в тилакоидной мембране. Фотосистемы тилакоидных мембран составлены из нескольких сотен антенных молекул хлорофиллов и вспомогательных пигментов, определенным образом ориентированных относительно фотохимического реакционного центра. Поглощение фотона одной из антенных молекул хлорофилла переводит эту молекулу в возбужденное состояние. Затем энергия возбуждения (экситон) быстро переносится по пигментным молекулам к реакционному центру (путь экситона обозначен черной стрелкой). Тилакоидные мембраны включают также цитохромный комплекс b6f и АТР-синтазу (см. рис. 19-60).
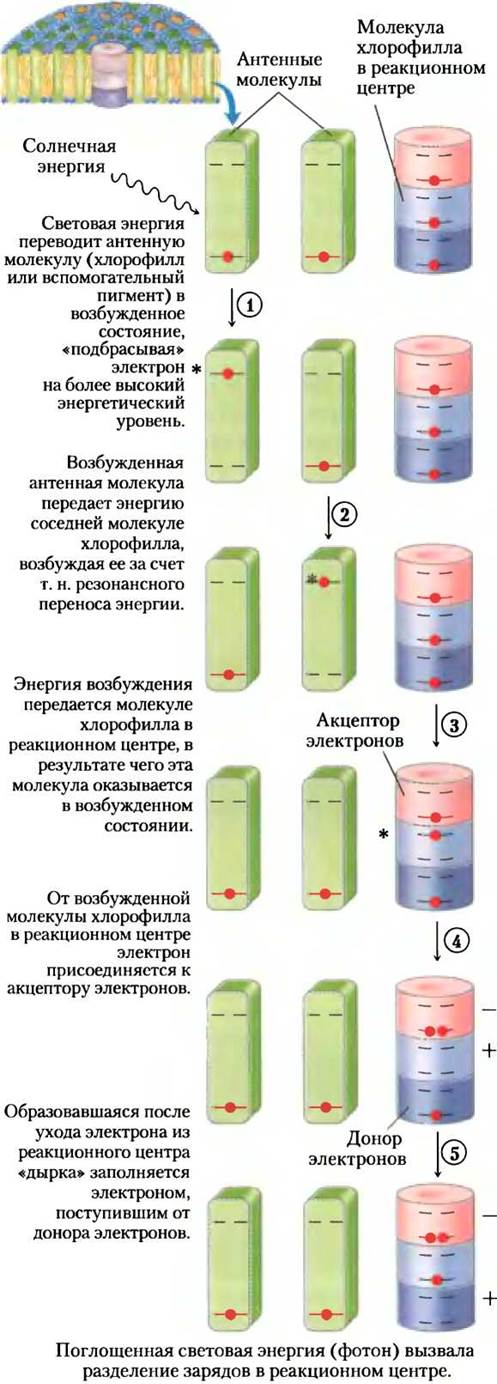
Светопоглощающие свойства молекул хлорофилла в фотохимических реакционных центрах и изолированных молекул сильно различаются. При освещении раствора хлорофилла in vitro поглощенная энергия быстро теряется в виде флуоресценции или рассеивается в виде тепла. Однако хлорофилл в интактных листьях под действием видимого света переходит в возбужденное состояние при очень небольшой интесивности флуоресценции (рис. 19-53, стадия (1)). Вместо этого в хлоропластах поэтапно происходят другие события. Возбужденная антенная молекула хлорофилла передает энергию непосредственно соседней молекуле, переводя ее в возбужденное состояние, а сама возвращается в основное состояние (стадия (2)). Затем энергия возбуждения (экситон) быстро мигрирует через третью, четвертую и т. д. молекулу пигмента к фотохимическому реакционному центру, в котором одна молекула хлорофилла в особой паре хлорофиллов типа а переходит в возбужденное состояние (стадия (3)). В возбужденной молекуле хлорофилла один из электронов переходит на более высокую энергетическую орбиталь. Этот возбужденный электрон покидает молекулу хлорофилла в реакционном центре и присоединяется к первому переносчику в цепи переноса электронов, оставляя в реакционном центре так называемую «дырку», обычно обозначаемую на схемах знаком «+» (рис. 19-53) (стадия (4)). Первый переносчик в цепи переноса электронов оказывается восстановленным, поскольку он присоединяет электрон, а реакционный центр — окисленным, поскольку он отдал свой электрон. Потеря электрона молекулой хлорофилла в реакционном центре восполняется электроном от соседней электронодонорной молекулы, приобретающей положительный заряд (стадия (5)). Таким образом, световая энергия, поглощаемая хлоропластами, индуцирует цепь окислительновосстановительных реакций.
Рис 19-53. Путь экситонов и электронов в хлоропластах. Преобразование поглощаемой энергии фотонов в транспорт зарядов в фотохимических комплексах хлоропластов осуществляется в несколько стадий. Стадия (1) повторяется в антенных молекулах до тех пор, пока экситон не достигнет молекулы хлорофилла в реакционном центре и не переведет ее в возбужденное состояние. О других состояниях см. текст. Звездочкой обозначено возбужденное состояние молекулы.
Краткое содержание раздела 19.7 Поглощение света
■ Фотофосфорилирование в хлоропластах зеленых растений и цианобактерий сопряжено с переносом электронов по цепи переносчиков.
■ В световых реакциях фотосинтеза зеленых растений поглощенная световая энергия возбуждает молекулы хлорофилла и вспомогательных пигментов, которые затем передают энергию возбуждения реакционным центрам в тилакоидных мембранах. В реакционных центрах фотовозбуждение создает электрические заряды, в результате чего образуется сильный донор электронов (восстановитель) и сильный акцептор электронов (окислитель).