ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
19. ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ И ФОТОФОСФОРИЛИРОВАНИЕ
Вопросы и задачи
1. Окислительно-восстановительные реакции.
NADH-дегидрогеназный комплекс митохондриальной дыхательной цепи катализирует следующие окислительно-восстановительные реакции:
(1) NADH + Н+ + E-FMN —> NAD++ E-FMNH2
(2) E-FMNH2 +2 Fe3+—> E-FMN + 2 Fe2+ + 2 H+
(3) 2 Fe2+ 2 H+ + Q —> 2 Fe3+ + QH2
Суммарная реакция:
NADH + H+ + Q —> NAD+ + QH2
Здесь Fe3+ и Fe2+ — атомы железа железосерных центров, Q — убихинон, QH2 — убихинол и Е — фермент.
Укажите для каждой из этих реакций, катализируемых NADH-дегидрогеназным комплексом: а) донор электронов, б) акцептор электронов, в) сопряженную окислительно-восстановительную пару, г) восстановитель, д) окислитель.
2. Все части молекулы убихинона выполняют определенную функцию.
При переносе электронов по митохондриальной дыхательной цепи только хиноновая часть убихинона претерпевает окислительно-восстановительные превращения, а боковая изопреноидная цепь остается без изменений. Какова роль этой цепи в процессах переноса электронов?
3. При окислении сукцината акцептором электронов служит не NAD+, a FAD.
Для всех дегидрогеназ, участвующих в процессах гликолиза и в цикле лимонной кислоты, акцептором электронов служит NAD+, Е'° (NAD+ / NADH) = -0,32 В. Единственное исключение — реакция, катализируемая сукцинатдегидрогеназой, использующей в качестве акцептора электронов ковалентно связанный с ней FAD; в комплексе с ферментом Е'° (FAD/ FADH2) = 0,050 В. Сравните Е'° для NAD+ / NADH и для FAD / FADH2 в комплексе с ферментом со стандартным восстановительным потенциалом сопряженной пары фумарат / сукцинат (Е'° = 0,031 В), и объясните, почему при дегидрировании сукцината FAD — более подходящий акцептор электронов, чем NAD+.
4. Степень восстановления переносчиков электронов в дыхательной цепи.
Степень восстановления каждого из переносчиков электронов в дыхательной цепи митохондрий определяется условиями, существующими в митохондриях. Если NADH и молекулярного кислорода достаточно, степень восстановления переносчиков электронов снижается при переходе электронов от субстрата на кислород. При блокировании переноса электронов переносчики, в дыхательной цепи занимающие место перед стадией блокирования, становятся более восстановленными, а те, которые располагаются после стадии блокирования, — более окисленными, как это показано на рис. 19-6. Укажите степени окисления убихинона, цитохромов b, с1, с и а + а3 для следующих четырех случаев:
а) достаточно NADH и O2, но добавлен цианид;
б) достаточно NADH, но исчерпан запас O2;
в) достаточно O2, но исчерпан запас NADH;
г) достаточно NADH и O2.
5. Влияние ротенона и антимицина А на перенос электронов.
Ротенон — токсичное вещество, вырабатываемое одним из видов растений, резко подавляет активность митохондриальной NADH- дегидрогеназы у насекомых и рыб. Токсичный антибиотик антимицин А сильно ингибирует окисление убихинола.
а) Почему ротенон оказывается смертельным ядом для некоторых насекомых и рыб?
б) Почему антимицин — яд?
в) Предположим, что оба эти вещества блокируют соответствующие участки дыхательной цепи с одинаковой эффективностью. Какое из них будет при этом более мощным ядом?
6. Разобщающие агенты при окислительном фосфорилировании.
В нормальных митохондриях скорость переноса электронов строго регулируется потребностями в АТР. Поэтому, если скорость использования АТР сравнительно невелика, небольшой оказывается и скорость переноса электронов. Если АТР расходуется с большой скоростью, скорость переноса электронов тоже высокая. В подобных условиях (при тесном сопряжении этих двух процессов) отношение Р/О, т. е. число образовавшихся молекул АТР в расчете на один атом потребленного
кислорода, когда донором электронов служит NADH, составляет приблизительно 2,5.
а) Как должна повлиять относительно низкая и относительно высокая концентрация разобщающего агента на скорость переноса электронов и на величину Р/О?
б) Прием внутрь разобщающих агентов вызывает обильное потоотделение и повышение температуры тела. Дайте объяснение этому феномену на молекулярном уровне. Как изменяется отношение Р/О в присутствии разобщающих агентов?
в) Разобщающий агент 2,4-динитрофенол пытались использовать для борьбы с ожирением. На чем может быть основано подобное действие этого агента? В настоящее время разобщающие агенты такого рода не применяются в качестве лекарственных препаратов, поскольку известны случаи, когда они приводили к летальному исходу. Почему прием разобщающих агентов может вызвать смерть?
7. Влияние валиномицина на окислительное фосфорилирование.
Если к суспензии активно дышащих митохондрий добавить антибиотик валиномицин, наблюдается уменьшение образования АТР и скорости поглощения кислорода, но происходит выделение тепла и увеличение трансмембранного градиента pH в митохондриях. Дайте объяснение этим фактам исходя из способности валиномицина переносить ионы К+ через внутреннюю мембрану митохондрий.
8. Механизм действия дициклогексилкарбодиимида (ДЦКД).
Если к суспензии активно дышащих митохондрий, в которых дыхание тесно сопряжено с фосфорилированием, добавить ДЦКД, наблюдается резкое уменьшение скорости переноса электронов (оценивают по количеству поглощенного кислорода) и скорости фосфорилирования (оценивают по образованию АТР). При добавлении к таким ингибированным митохондриальным препаратам 2,4-динитрофенола потребление кислорода возвращается к норме, но синтез АТР так и остается подавленным.
а) На какую стадию процесса переноса электронов или окислительного фосфорилирования влияет ДКЦД?
б) Почему ДКЦД нарушает потребление кислорода в митохондриях? Каков механизм действия 2,4-динитрофенола на препарат ингибированных митохондрий?
в) С каким из перечисленных ниже ингибиторов более всего сходен по своему действию ДКЦД: с антимицином А, ротеноном, олигомицином или арсенатом?
9. Компартментация компонентов цикла лимонной кислоты.
Обнаружено, что в окислительновосстановительных реакциях изоцитратдегидрогеназа находится только в митохондриях, а малат- дегидрогеназа — и в цитозоле, и в митохондриях. Какова роль цитозольной малатдегидрогеназы в цикле лимонной кислоты?
10. Малат-α-кетоглутаратная транспортная система.
Транспортная система, осуществляющая перенос малата и α-кетоглутарата через внутреннюю мембрану (см. рис. 19-27), блокируется н-бутилмалонатом. К аэробной суспензии клеток почек, использующих в качестве «топлива» только глюкозу, добавляют н-бутилмалонат. Какое влияние окажет этот ингибитор на процессы в клетках почки? А именно на следующие процессы:
а) гликолиз;
б) поглощение кислорода;
в) образование лактата;
г) синтез АТР.
11. Синтез АТР регулируется концентрацией ADP в клетках.
Для синтеза АТР путем окислительного фосфорилирования требуются ADP и фосфат, но скорость синтеза зависит главным образом от концентрации ADP в клетках. Дайте аргументированное объяснение этому факту.
12. Временная шкала в регуляторных процессах в митохондриях.
Сравните между собой промежутки времени, необходимые для регуляции скорости дыхания, при (а) повышении концентрации ADP и (б) снижении рO2. В чем причина различий?
13. Эффект Пастера.
Если в суспензию анаэробных клеток, потребляющих глюкозу с большой скоростью, ввести кислород, клетки начнут его поглощать и уровень потребления глюкозы резко понизится. Одновременно с этим прекратится на копление лактата. Этот эффект, характерный для клеток, способных и к аэробному, и к анаэробному потреблению глюкозы, впервые наблюдал Луи Пастер в 60-х гг. XIX в., и потому он был назван эффектом Пастера.
а) Почему при введении в клеточную суспензию кислорода прекращается накопление лактата?
б) Почему в присутствии кислорода снижается скорость потребления глюкозы?
в) Каким образом после начавшегося потребления кислорода понижается скорость потребления глюкозы? Объясните это на основе специфического действия ферментов.
14. Мутанты дрожжей с нарушением процесса дыхания клеток и образование спирта.
Мутанты дрожжей с нарушением способности потреблять кислород (р-, «petites») получены воздействием мутантных агентов на нормальные клетки дрожжей. Такие мутантные клетки дрожжей не способны синтезировать фермент цитохромоксидазу, в результате чего вызванные ими процессы брожения не подавляются кислородом, т. е. в них не проявляется эффект Пастера (см. задачу 13). Некоторые производители спирта используют мутантные дрожжи для получения спирта в больших количествах из древесных опилок. Объясните преимущества использования для этого мутантных дрожжей по сравнению с нормальными дрожжевыми клетками. Почему при отсутствии в клетках фермента цитохромоксидазы эффект Пастера не наблюдается?
15. Преимущества суперкомплексов при передаче электронов.
Появляются новые доказательства того, что митохондриальные комплексы I, II, III и IV являются частью более крупного суперкомплекса. Каким преимуществом может обладать система, в которой все четыре комплекса собраны в единый суперкомплекс?
16. Сколько ионов Н+ содержится в одной митохондрии.
Хемиосмотическая гипотеза предполагает, что в результате переноса электронов ионы Н+ «выталкиваются» из матрикса митохондрий наружу, вследствие чего между двумя сторонами митохондриальной мембраны возникает градиент pH, при котором наружная сторона оказывается более кислой, чем внутренняя. Способность ионов Н+диффундировать в обратном направлении — из окружающей среды в митохондриальный матрикс (где их концентрация ниже) — служит движущей силой синтеза АТР, катализируемого ATP-синтазой. В митохондриях, суспендированных в среде с pH 7,4, происходит окислительное фосфорилирование. Показано, что митохондриальный матрикс имеет pH 7,7.
а) Вычислите молярные концентрации ионов Н+ в окружающей среде и в матриксе для этих условий.
б) Определите отношение концентраций Н+ снаружи и внутри, что поможет оценить энергию, которая потенциально соответствует этой разности концентраций (см. уравнение 11-4, т. 1).
в) Определите число ионов Н+, приходящихся на одну дышащую митохондрию печени. При расчетах примите, что внутреннее пространство митохондрии представляет собой сферу диаметром 1,5 мкм.
г) Учитывая результаты ваших расчетов, ответьте, можно ли считать, что достаточно только одного этого градиента pH как источника энергии для синтеза АТР?
д) Если одного градиента pH не достаточно, то какой другой источник энергии, необходимой для синтеза АТР, вы могли бы указать?
17. Скорость превращения АТР в сердечной мышце крысы.
Аэробные клетки сердечной мышцы крысы получают более 90% необходимого им количества АТР в процессе окислительного фосфорилирования, используя в качестве «топлива» глюкозу. Каждый грамм ткани сердечной мышцы крысы потребляет кислород со скоростью 10,0 мкмоль/мин.
а) Рассчитайте скорость потребления глюкозы и образования АТР в сердечной мышце крысы.
б) Рассчитайте, на сколько секунд хватит запаса АТР в клетке, если начальная концентрация АТР была равна 5,0 мкмоль на 1 г мышечной ткани. Что означает полученный результат, учитывая необходимость точного регулирования образования АТР в клетках? При расчетах принимайте во внимание, что концентрация выражается в мкмоль/г мышечной ткани (эта ткань состоит в основном из воды).
18. Скорость расщепления АТР в летательных мышцах.
В летательных мышцах зеленой падальной мухи (Lucilia sericata) АТР образуется только в процессе окислительного фосфорилирования. Для поддержания во время полета концентрации АТР 7,0 мкмоль на 1 г мышцы насекомое должно потреблять кислород со скоростью 187 мл /ч на грамм своей массы. Рассчитайте скорость расходования АТР в летательных мышцах, если эти мышцы составляют 20% массы насекомого. В течение какого времени пул АТР может не пополняться за счет окислительного фосфорилирования? Предполагается, что восстановительные эквиваленты переносятся глицерин-3-фосфатной транспортной системой и что потребляемый кислород находится под давлением 101,3 кПа (1 атм) при температуре 25 °С.
19. Дефекты митохондрий и рак.
Мутации генов, кодирующих некоторые митохондриальные белки, связаны с повышением риска развития некоторых типов рака. Как дефекты митохондрий могут приводить к раку?
20. Вариабельность тяжести митохондриальных заболеваний.
У людей с заболеваниями, вызванными определенным дефектом митохондриального генома, симптомы заболевания могут носить как умеренный, так и сильно выраженный характер. Объясните этот факт.
21. Перенос восстановительных эквивалентов через мембрану.
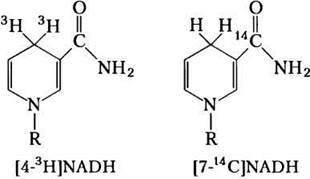
В аэробных клетках внемитохондриальный NADH должен окисляться в дыхательных цепях митохондрий. При введении NADH, меченного радиоактивным изотопом водорода (4-3H[NADH]) в препарат гепатоцитов крысы, содержащих митохондрии и все цитозольные ферменты, радиоактивность вскоре обнаруживалась в митохондриальном матриксе. При введении 7-14C[NADH] матрикс оставался нерадиоактивным. Какие выводы о процессах окисления внемитохондриального NADH позволяют сделать эти наблюдения?
22. Высокий уровень аланина в крови связан с нарушением окислительного фосфорилирования.
Выяснилось, что у большинства людей с нарушениями окислительного фосфорилирования концентрация аланина в крови довольно высокая. Объясните это наблюдение.
23. Функции запасенных NAD и дегидрогеназы.
Для пируватдегидрогеназы и глицеральдегид-3- фосфатдегидрогеназы акцептором электронов служит NAD+, хотя оба эти фермента локализуются в различных структурах эукариотических клеток. Объясните этот факт.
24. Диабет как следствие митохондриальных дефектов.
Глюкокиназы играют важную роль в метаболизме глюкозы в β-клетках поджелудочной железы. У людей с двумя дефектными копиями гена глюкокиназы проявляется тяжелый врожденный диабет, а при дефектах только одной копии развивается заболевание гораздо меньшей степени тяжести (диабет зрелого типа у молодых, MODY2). Объясните это различие, учитывая свойства β-клеток.
25. Последствия мутаций в митохондриальном комплексе II.
Замена одного-единственного нуклеотида в гене сукцинатдегидрогеназы (комплекс II) связана с возникновением карциномы средней части тонкого кишечника. Предложите механизм, объясняющий это наблюдение.
26. Фотохимическая эффективность света с различной длиной волны.
При освещении зеленого растения светом с длиной волны 680 или 700 нм скорость фотосинтеза, измеряемая по выделению кислорода O2, в первом случае оказывается выше. Однако одновременное освещение растения светом той и другой длины волны обеспечивает более высокую скорость фотосинтеза, чем освещение светом каждой длины волны. Объясните причину этого.
27. Расчет баланса фотосинтеза.
В 1804 г. Теодор Соссюр показал, что суммарная масса кислорода, выделяемого растением в процессе фотосинтеза, и синтезированного вещества в сухом виде больше массы поглощенного углекислого газа. Объясните этот факт.
28. Роль H2S у некоторых фотосинтезирующих бактерий.
У пурпурных серных бактерий на свету в присутствии Н2O и 14СO2 может идти фотосинтез, но только если имеется H2S, а кислород отсутствует. В ходе фотосинтеза (скорость которого измеряют по образованию 14С-глюкозы) H2S превращается в элементную серу, а кислород не выделяется. Какую роль играет превращение H2S в элементную серу? Почему не выделяется кислород?
29. Увеличение восстановительной силы фотосистемы I за счет энергии света.
При поглощении фотосистемой I красного света с длиной волны 700 нм стандартный потенциал восстановления ее реакционного центра Р700 меняется от 0,40 до -1,2 В. Какая доля поглощенного света запасается как восстановительная сила?
30. Поток электронов в фотосистемах I и II.
Предскажите, каким образом ингибитор прохождения электронов через феофитин повлияет на перенос электронов через (а) ФСI и (б) ФСII. Объясните свои рассуждения.
31. Ограниченный синтез АТР в темноте.
Если хлоропласты шпината освещать в отсутствие АТР и фосфата, а затем добавить ADP и фосфат в темноте — в течение короткого промежутка времени наблюдается синтез АТР. Объясните результаты этого эксперимента.
32. Механизм действия гербицида диурона.
Если хлоропласты обработать таким гербицидом, как диурон, выделение кислорода и фотофосфорилирование прекращаются. Выделение кислорода (но не фотофосфорилирование) можно восстановить, добавив какой-нибудь внешний акцептор электронов, например, реагент Хилла. Как действует этот гербицид, когда он убивает сорняки? В какой точке схемы, изображенной на рис. 19-56, может сказываться ингибирующее действие диурона? Дайте аргументированный ответ.
33. Влияние вентурицидина на превращения кислорода.
Вентурицидин — мощный ингибитор АТР-синтазы; он взаимодействует с CF0- компонентом фермента и блокирует прохождение протонов через комплекс CF0CF1. Как вентурицидин повлияет на превращения кислорода в суспензии хорошо освещенных хлоропластов? Произойдут ли изменения, если эксперимент проводить в присутствии разобщающего агента, такого как 2,4-динитрофенол? Объясните свой ответ.
34. Биоэнергетика фотофосфорилирования.
Стационарные концентрации АТР, ADP и фосфата в изолированных листьях шпината при полном освещении и pH 7 равны 120,0, 6,0 и 700 мкМ соответственно.
а) Сколько требуется свободной энергии для синтеза 1 моль АТР при этих условиях?
б) Энергию для синтеза АТР поставляет перенос электронов в хлоропластах, индуцированный светом. При какой минимальной разности потенциалов должен происходить перенос пары электронов в этих условиях для обеспечения синтеза АТР? (Можете воспользоваться уравнением 13-6, с. 47.)
35. Энергия света индуцирует окислительновосстановительные реакции.
Предположим, гипотетический фотосинтезирующий микроорганизм окисляет H2S и переносит электроны к NAD+. Какова должна быть длина волны поглощаемого света, чтобы его энергии хватило на отщепление электрона от H2S и передачу его NAD+ в стандартных условиях? Эффективность фотохимического процесса в рассматриваемом случае составляет 100%, а стандартные восстановительные потенциалы для H2S и NAD+ равны -243 и -320 мкВ соответственно. Примите во внимание схему на рис. 19-46.
36. Константы равновесия реакций расщепления воды.
Кофермент NADP+ — конечный акцептор электронов в хлоропластах, что можно записать в виде уравнения реакции:
2 Н2O + 2 NADP+ —> 2 NADPH + 2 Н+ + O2
Используя данные табл. 19-2, рассчитайте константы равновесия этой реакции при 25 °С. (Соотношения между К'eq и ∆G'° обсуждаются на с. 15.) За счет какой энергии протекает эта реакция в хлоропластах?
37. Энергетика фотопреобразований.
В процессе фотосинтеза растений образование одной молекулы O2 требует поглощения восьми квантов света.
2 Н2O + 2 NADP+ + 8 фотонов —> 2 NADPH + 2 Н+ + O2
Рассчитайте для этой реакции изменение свободной энергии, если длина волны поглощаемого света 700 нм (красный) и эффективность фотосинтезирующего процесса 100%.
38. Перенос электронов к реагенту Хилла.
Если к экстрактам хлоропластов шпината добавить феррицианид калия (реагент Хилла), а затем осветить эти препараты, будет выделяться кислород согласно уравнению
2 Н2O + 4 Fe3+ —> O2 + 4 Н+ + 4 Fe2+
где Fe3+ — феррицианид, Fe2+ — ферроцианид. Образуется ли в ходе этой реакции NADPH? Аргументируйте ответ.
39. Как часто молекула хлорофилла поглощает фотон?
В листьях шпината хлорофилл (Мr = 892) содержится в количестве 20 мкг/см2. В дневное время при средней освещенности 5,4 Дж/(см2 • мин) листья поглощают около 50% солнечной энергии. Как часто одна молекула хлорофилла поглощает фотон? Рассчитайте долю молекул хлорофилла, одновременно переходящих в возбужденное состояние, если среднее время жизни возбужденной молекулы хлорофилла in vivo 1 нс.
40. Влияние монохроматического света на поток электронов.
Степень окисления или восстановления переносчика электронов при фотосинтетическом переносе электронов в некоторых случаях можно определить из спектрофотометрических данных. Когда хлоропласты освещают светом с длиной волны 700 нм, цитохром f, пластоцианин и пластохинон окисляются. Однако если освещать хлоропласты светом с длиной волны 680 нм, эти переносчики электронов оказываются восстановленными. Объясните этот факт.
41. Функция циклического фотофосфорилирования.
Когда отношение концентраций [NADPH] / [NADP+] в хлоропласте высокое, происходит в основном циклическое фотофосфорилирование (см. рис. 19-56). Участвует ли кислород в циклическом фотофосфорилировании? Образуется ли NADPH. Аргументируйте ответ. Какова главная функция циклического фотофосфорилирования?
Анализ экспериментальных данных
42. Фотофосфорилирование: открытие, отклонение гипотезы и повторное открытие.
В 1930- 1940-х гг. ученые начали понимать механизм процесса фотосинтеза. Тогда же стала проясняться роль «энергетически богатых фосфатных связей» (сегодня это АТР) в гликолизе и клеточном дыхании. Существовало много теорий, объяснявших механизм фотосинтеза, особенно это касалось роли света. Данная задача рассматривает процесс, который тогда называли «первичным фотохимическим процессом», т. е. первичное превращение энергии поглощенного света в фотосинтезирующей клетке. Интересно, что одна важная составная часть современной модели фотосинтеза была сформулирована достаточно рано, но потом отклонена, забыта на несколько лет и лишь затем принята вновь.
В 1944 г. Эмерсон, Стауффер и Умбрейт предположили, что «функция световой энергии в фотосинтезе заключается в образовании «энергетически богатых» фосфатных связей» (с. 107). В соответствии с их моделью (далее мы будем называть ее моделью Эмерсона) свободная энергия, необходимая как для фиксации, так и для восстановления СO2, берется как раз из этих «энергетически богатых» фосфатных связей (т. е. из АТР), образующихся в результате поглощения света хлорофиллсодержащим белком.
Эта модель была опровергнута Рабиновичем (1945). Суммируя открытия Эмерсона с соавт., Рабинович заключил: «Пока не получено других данных, мы выдвигаем против данной гипотезы аргумент, вытекающий из энергетических соображений. Фотосинтез представляет собой процесс накопления энергии. Зачем нужно превращать квант света (даже красного света, энергия которого около 43 ккал/эйнштейн) в «фосфатный квант» с энергией всего лишь 10 ккал/моль? Это был бы шаг в ложном направлении — не к накоплению, а к рассеиванию энергии» (т. 1, с. 228). Этот аргумент, а также другие наблюдения привели к отклонению модели Эмерсона. Только в 1950-х гг. модель Эмерсона была признана верной, хотя и с некоторыми модификациями.
«Пройдитесь» по информации (см. ниже пункты а-ж), изложенной в статье Эмерсона с соавторами. Ответьте на три вопроса:
1. Каким образом эта информация обосновывает модель Эмерсона, в соответствии с которой энергия света напрямую используется хлорофиллом для синтеза АТР, а затем АТР обеспечивает энергию, необходимую для фиксации и восстановления СO2?
2. Как бы мог Рабинович объяснить эту информацию на основании своей модели (и большинства существовавших тогда моделей), в которой световая энергия использовалась хлорофиллом напрямую для синтеза восстанавливающих соединений? Рабинович писал: «Теоретически, нет причин, по которым вся энергия электронов, содержащаяся в молекулах, возбужденных в результате поглощения света, не была бы доступна для процессов окисления и восстановления» (т. 1, с. 152). В этой модели восстанавливающие вещества далее использовались для фиксации СO2, и его восстановления, а необходимая для этого энергия составляла всего лишь некоторую долю того большого количества свободной энергии, которое образуется при реакциях восстановления.
3. Как данная информация объясняется в свете наших современных представлений о процессе фотосинтеза?
а) Хлорофилл содержит ион Mg2+, который, как известно, является важным кофактором во многих ферментах, катализирующих реакции фосфорилирования и дефосфорилирования.
б) Грубый препарат «хлорофиллового белка», выделенный из фотосинтезирующих клеток, обладал фосфорилирующей активностью.
в) Фосфорилирующая активность «хлорофиллового белка» ингибировалась светом.
г) Уровень содержания некоторых фосфорилированных соединений в фотосинтезирующих клетках сильно изменялся на свету. (Эмерсону с соавт. не удалось идентифицировать соответствующие вещества).
Как выяснилось позднее, в какой-то степени были верны обе модели — и Эмерсона, и Рабиновича.
д) Объясните, каким образом эти две модели соотносятся с современной моделью фотосинтеза.
При опровержении модели Эмерсона Рабинович настаивал на следующем: «Проблема теории запасания фосфата наиболее отчетливо проявляется тогда, когда мы обратимся к тому факту, что при слабом освещении восьми или десяти квантов света достаточно для восстановления одной молекулы диоксида углерода. Если каждый квант должен произвести одну молекулу высокоэнергетического фосфата, будет накоплено лишь 80-100 ккал/эйнштейн, тогда как для фотосинтеза необходимо не менее 112 ккал/моль или более — ввиду потерь в необратимых частичных реакциях» (т. 1, с. 228).
е) Как расчеты Рабиновича (от 8 до 10 фотонов на молекулу восстановленного СO2) соотносятся с принятым сегодня значением? За некоторой дополнительной информацией обратитесь к гл. 20.
ж) Как бы вы отклонили аргумент Рабиновича на основании наших сегодняшних знаний о процессе фотосинтеза?