ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 2. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ - 2014
ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
20. БИОСИНТЕЗ УГЛЕВОДОВ У РАСТЕНИЙ И БАКТЕРИЙ
20.2. Фотодыхание, С4- и САМ-пути
Как мы видели, в фотосинтезирующих клетках в световых реакциях (гл. 19) образуется O2 (из Н2O), а в светонезависимых реакциях потребляется СО2, поэтому при фотосинтезе происходит изменение газового состава окружающей среды — поглощение СO2 и высвобождение O2.
СO2 + Н2O —> O2 + (СН2O)
В темноте растения осуществляют также митохондриальное дыхание — окисление субстратов до СO2 и восстановление O2 до Н2O. Однако у растений происходит и другой процесс, при котором, как и при митохондриальном дыхании, потребляется O2 и выделяется СO2, и который, как фотосинтез, управляется светом. Этот процесс фотодыхания — дорогостоящая реакция фотосинтеза по причине недостаточной специфичности фермента рубиско. В следующем разделе мы опишем эту побочную реакцию и стратегии, которые используются растениями для минимизации ее метаболических последствий.
Фотодыхание — следствие оксигеназной активности рубиско
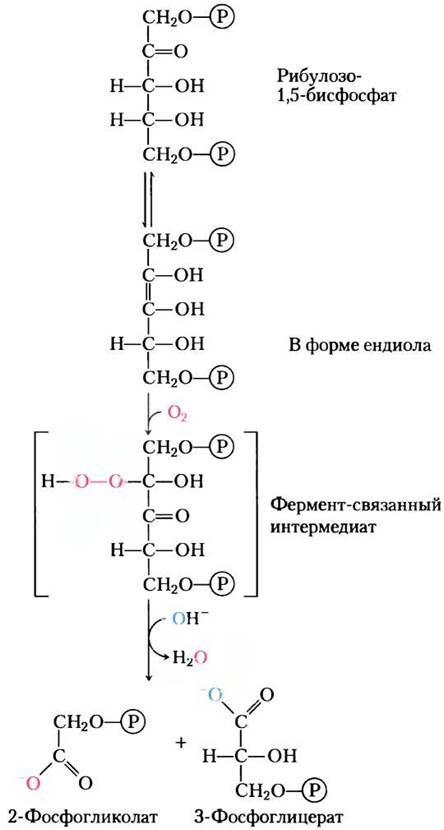
Рубиско не имеет абсолютной специфичности в отношении СO2 как субстрата. Молекулярный кислород (O2) конкурирует с СO2 за активный центр, и через каждые три-четыре оборота рубиско катализирует одну конденсацию O2 с рибулозо-1,5-бисфосфатом с образованием 3-фосфоглицерата и 2-фосфогликолата (рис. 20-20), метаболически бесполезного продукта. В этом и проявляется оксигеназная активность этого фермента, которая отражена в его полном названии: рибулозо-1,5-бисфосфат- карбоксилаза/оксигеназа. В результате реакции с O2 не происходит фиксации углерода, и в итоге такая реакция оказывается обузой для клетки; усвоение атомов углерода из 2-фосфогликолата требует от клетки много энергии и высвобождает некоторое количество ранее фиксированного углекислого газа.
Если реакция с кислородом вредна для организма, то почему же в ходе эволюции активный центр рубиско не «научился» различать O2 и СO2? Возможно, большинство эволюционных изменений фермента произошла около 2,5 млрд лет назад, еще до того времени, как фотосинтезирующие организмы начали повышать содержание кислорода в атмосфере, т. е. это было тогда, когда рубиско не требовалось различать O2 и СO2. Для СO2 константа Михаэлиса Км ≈ 9 мкМ, для О2 Км ≈ 350 мкМ. В нашей атмосфере около 20% O2 и только 0,04% СO2, в воде, находящейся в равновесии с атмосферным воздухом при комнатной температуре, концентрации этих газов составляют -250 мкМ O2 и -11 мкМ СO2; такие концентрации способствуют «фиксации» O2 с помощью рубиско и, следовательно, вызывает значительные затраты
энергии. Температурная зависимость относительной растворимости O2 и СO2 ([O2] / [СO2]) круто возрастает при высоких температурах. Кроме того, сродство рубиско к СO2уменьшается с увеличением температуры, усиливая вероятность катализа расточительной оксигеназной реакции. А так как СO2 потребляется в реакциях ассимиляции, соотношение между O2 и СO2 в воздушном пространстве, которое окружает листья, увеличивается, что еще больше способствует оксигеназной реакции.
Рис. 20-20. Оисигеназная активность рубиско. Рубиско может включать O2 вместо СO2 в рибулозо-1,5-бисфосфат. Образующийся нестабильный интермедиат распадается на 2-фосфогликолат (он перерабатывается, как показано на рис. 20-21) и 3-фосфоглицерат, который может включаться в цикл Кальвина.
Усвоение фосфоглицерата обходится клетке весьма дорого
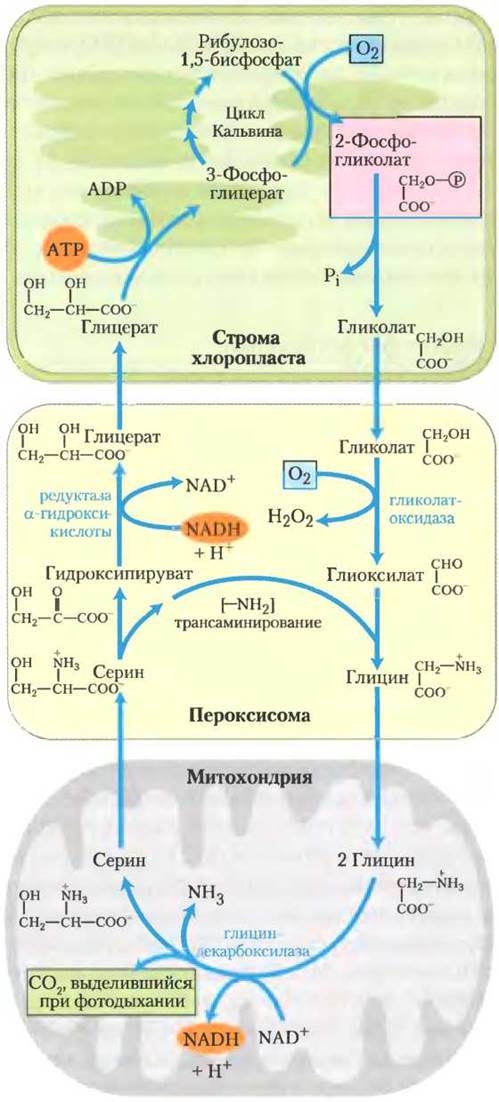
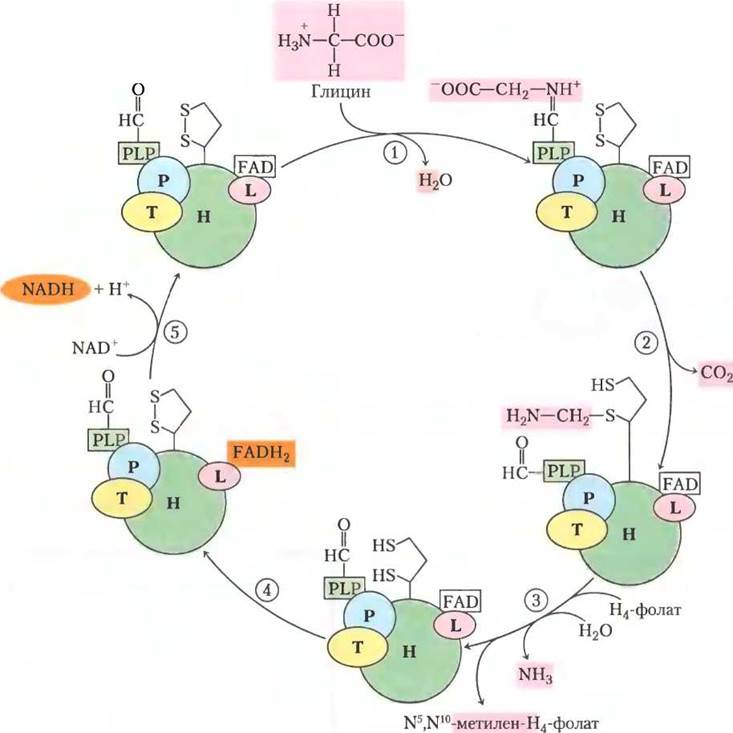
Гликолатный путь осуществляет превращение двух молекул 2-фосфогликолата в молекулу серина (3 атома углерода) и молекулу СO2 (рис. 20-21). В хлоропласте фосфатаза переводит 2-фосфогликолат в гликолат, который экспортируется в пероксисому. Там гликолат окисляется молекулярным кислородом, и получившийся альдегид (глиоксилат) подвергается трансаминированию с образованием глицина. В качестве побочного продукта реакции окисления гликолата образуется пероксид водорода, который распадается под действием пероксидазы пероксисомы. Глицин транспортируется из пероксисомы в митохондрию, где подвергается окислительному декарбоксилированию под действием глициндекарбоксилазного комплекса. Этот фермент похож по структуре и механизму действия на митохондриальные ферменты, с которыми мы уже сталкивались: пируватдегидрогеназный и α-кетоглутаратдегидрогеназный комплексы (гл. 16). Глициндекарбоксилазный комплекс окисляет глицин до СO2 и NН3 с одновременным восстановлением NАD+ до NАDН и переносом оставшегося атома углерода от глицина на кофактор тетрагидрофолат (рис. 20-22). Получившаяся одноуглеродная единица затем переносится серин-гидроксиметилтрансферазой, синтезирующей серин, с тетрагидрофолата на второй глицин. Итоговая реакция, катализируемая глициндекарбоксилазным комплексом и серин- гидроксиметилтрансферазой приведена ниже.
2 Глицин + NАD+ + Н2O —> серин + СO2 + NН3 + NАDН + Н+
Затем серин превращается в гидроксипируват, далее в глицерат и в конце концов в 3-фосфо- глицерат, который используется для регенерации рибулозо-1,5-бисфосфата, завершая этот длинный «дорогой» цикл (рис. 20-21).
Рис. 20-21. Гликолатный путь. Путь усвоения 2-фос- фоглицерата (выделен розовым) через его превращение в серин и в конечном счете в 3-фосфоглицерат, в котором участвуют три компартмента клетки. Гликолат, образующийся при дефосфорилировании 2-фосфогликолата в хлоропластах, окисляется в пероксисомах до глиоксилата и затем трансаминируется до глицина. В митохондриях две молекулы глицина конденсируются с образованием серина, и СO2 высвобождается в процессе фотодыхания (выделен зеленым). Эта реакция катализируется глициндекарбоксилазой, ферментом, присутствующим в высоких концентрациях в митохондриях С3-растений (см. текст). Серин превращается в гидроксипируват и затем в глицерат в пероксисомах; глицерат попадает в хлоропласты, фосфорилируется и включается в цикл Кальвина. Кислород (окрашен голубым) потребляется в процессе фотодыхания в две стадии.
При ярком солнечном свете утечка через гликолатный путь может быть очень высокой. Растение производит таким способом в 5 раз больше СO2, чем обычно образуется в результате всех окислительных реакций в цикле трикарбоновых кислот. Для обеспечения этого потока митохондрии растений содержат потрясающе огромное количество глициндекарбоксилазного комплекса: в листьях гороха и шпината четыре белка этого комплекса составляют половину всех белков митохондриального матрикса! В нефотосинтезирующих частях растения, таких как картофельные клубни, митохондрии содержат глициндекарбоксилазный комплекс в очень низкой концентрации.
Совместная активность рубиско-оксигеназы и гликолатного пути потребляет O2 и производит СO2 — откуда и название фотодыхание. Возможно, этот путь лучше назвать цикл фотосинтетического окисления углерода или С2-путь; эти названия не вызывают ассоциации с дыханием в митохондрии. В отличие от митохондриального дыхания фотодыхание не сохраняет энергию и может ингибировать образование чистой биомассы почти на 50%. Эта неэффективность ведет к эволюционным адаптациям, особенно у растений, которые развиваются в теплом климате.
У С4-растений фиксация СO2 и активность рубиско пространственно разделены
У многих растений, которые растут в тропиках, а также теплолюбивых культур, происходящих из тропиков, таких как кукуруза, сахарный тростник и сорго, развился механизм, благодаря которому они избегают проблемы невыгодного фотодыхания. Стадии включения СO2 в трехуглеродный 3-фосфоглицерат предшествует несколько стадий, в том числе временная фиксация СO2 в четырехуглеродное соединение. Растения, использующие этот путь, называются С4-растениями, а сам процесс ассимиляции — С4-метаболизмом, или С4- путем. Растения, которые фиксируют СO2 по пути, где на первой стадии происходит реакция конденсация СO2 с рибулозо-1,5-бисфосфатом с образованием 3-фосфоглицерата, называются С3-растениями.
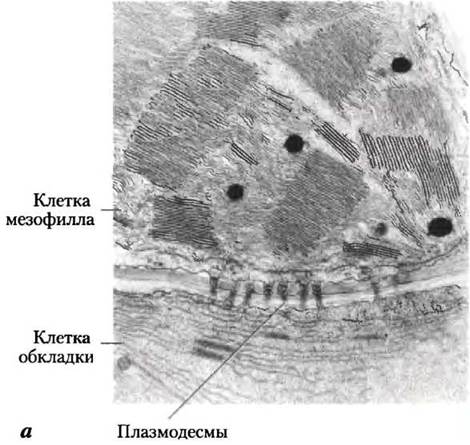
С4-Растения обычно развиваются при большой интенсивности света и высокой температуре. Их отличает несколько важных характеристик: высокая скорость фотосинтеза, большая скорость роста, низкий уровень фотодыхания, малая скорость потери влаги и специализированная структура листа. В листьях С4-растений в фотосинтезе участвуют два типа клеток: мезофилл и клетки обкладки (рис. 20-23, а). Существуют три варианта С4-метаболизма, расшифрованные в 1960-х гг. Маршаллом Хэтчем и Роджером Слэком (рис. 20-23, б).
Рис. 20-22. Глициндекарбоксилазная система. Глициндекарбоксилаза митохондрий растений представляет собой комплекс из четырех типов субъединиц со стехиометрией P4H27T9L2. С белком Н ковалентно связан остаток липоевой кислоты, которая может подвергаться обратимому окислению. (1) — образование шиффова основания между пиридоксальфосфатом (PLP от англ. pyridoxal phosphate) и глицином, катализируемое белком Р (названным так потому, что он образует связь с PLP). (2) — белок Р катализирует окислительное декарбоксилирование глицина, высвобождая СО2; оставшаяся группа метиламина связана с одной из -SH-групп восстановленной липоевой кислоты. (3) — белок Т (который использует тетрагидрофолат (H4F) в качестве кофактора) высвобождает NH3 из метиламина и переносит оставшийся одноуглеродный фрагмент на тетрагидрофолат, образуя N5, N10-метилентетрагидрофолат. (4) — белок L окисляет две -SH- группы липоевой кислоты до дисульфидной связи, (5) передавая электроны через FAD на NAD+ и завершая таким образом цикл. N5, N10-Метилентетрагидрофолат, образованный в этом процессе, используется серин-гидроксиметилтрансферазой для превращения молекулы глицина в серин, в результате регенерируется тетрагидрофолат, необходимый для реакции, катализируемой белком Т. L-Субъединица глициндекарбоксилазы идентична дигидролипоилдегидрогеназе (Е3) пируватдегидрогеназы и α-кетоглутаратдегидрогеназы (см. рис. 16-6).
У тропических растений первый интермедиат, в который включается 14СО2, — четырехуглеродное соединение оксалоацетат. Эта реакция происходит в цитозоле клеток мезофилла листьев и катализируется фосфоенолпируват- карбоксилазой, субстратом для которой служит НСО3-, а не СО2. Затем оксалоацетат либо восстанавливается до малата с расходом NADPH (см. рис. 20-26, б), либо превращается в аспартат в реакции трансаминирования:
Оксалоацетат + α-аминокислота —> L-аспартат + α-кетокислота
Малат или аспартат, синтезированный в клетках мезофилла, проникает в соседние клетки обкладки через плазмодесмы — образованные белками каналы, которые соединяют две растительные клетки и осуществляют движение метаболитов и даже малых белков между клетками. В клетках обкладки малат окисляется и декарбоксил ируется с образованием пирувата и СО2 под действием NADP-зависимой декарбоксилирующей малатдегидрогеназы (иногда называемой малик-ферментом, или яблочным ферментом) с восстановлением NADP+. У растений, использующих аспартат в качестве переносчика СО2, аспартат, поступивший в клетки обкладки, трансаминируется с образованием оксалоацетата и восстанавливается до малата, после чего СО2 высвобождается малатдегидро- геназой или ФЕП-карбоксикиназой. Как показали эксперименты с образцами, меченными 14С, молекула СО2, высвобождающаяся в клетках обкладки, — это та же самая молекула СО2, которая изначально находилась в оксалоаце- тате в клетках мезофилла. Эта молекула СО2 снова фиксируется, теперь с помощью рубиско, в той же самой реакции, которая происходит у С3-растениях: происходит включение СО2в положение С-1 молекулы 3-фосфоглицерата.
Пируват, образовавшийся при декарбокси- лировании малата в клетках обкладки, транспортируется обратно, в клетки мезофилла, где превращается в фосфоенолпируват в необычной реакции, катализируемой пируват-фосфатдикиназой (рис. 20-23, б). Этот фермент называется дикиназой, потому что он, используя одну молекулу АТР, одновременно фосфорилирует две разные молекулы: пируват до фосфоенолпирувата и фосфат до пирофосфата. Впоследствии пирофосфат гидролизуется до фосфата, поэтому получается, что для регенерации фосфоенолпирувата используются две высокоэнергетические фосфатные группы АТР. Теперь этот фосфоенолпируват готов для включения следующей молекулы СО2 в клетки мезофилла.
Рис. 20-23. Ассимиляция углерода у С4-растений. В растениях тропических районов преобладает С4-путъ, проходящий в клетках мезофилла и клетках обкладки, а — электронная микрофотография, показывающая хлоропласта мезофилла и прилегающих клеток обкладки. Клетки обкладки содержат гранулы крахмала. Видны плазмодесмы, соединяющие две клетки. б — С4-путь ассимиляции СO2, который происходит с участием четырехуглеродного интермедиата.
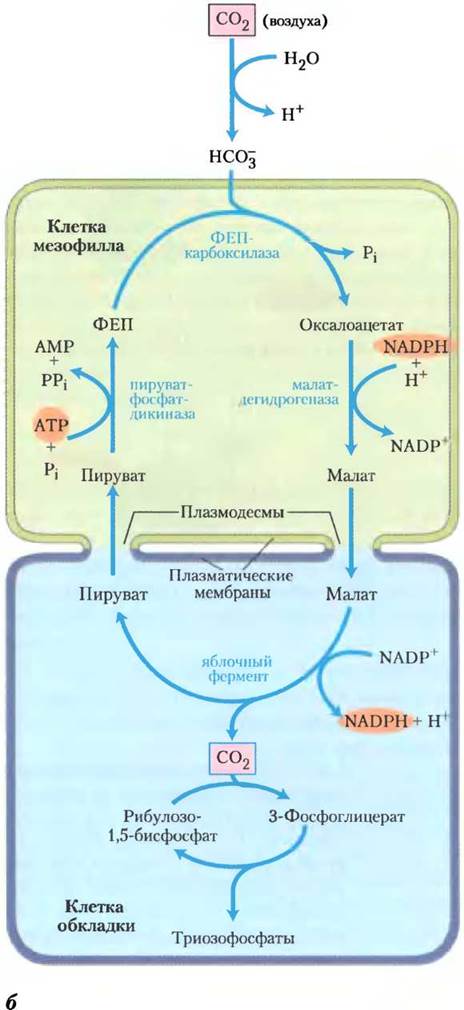
ФЕП-карбоксилаза имеет высокое сродство к НСО3- (который в водном окружении образуется из СО2) и фиксирует СО2 более эффективно, чем рубиско. В отличие от рубиско этот фермент не использует О2 в качестве альтернативного субстрата, поэтому в этой реакции нет конкуренции между СО2 и О2. Таким образом, ФЕП- карбоксилазная реакция служит для фиксации СО2 в форме малата. Высвобождением СО2 из малата в клетках обкладки достигается высокая локальная концентрация углекислого газа для функционирования рубиско с почти максимальной скоростью и подавления ее оксигеназной активности.
В клетках обкладки СО2 включается в 3-фосфоглицерат, поэтому другие реакции цикла Кальвина происходят так, как было описано ранее. Таким образом, у С4-растений клетки мезофилла осуществляют ассимиляцию СО2 по С4- пути, а клетки обкладки синтезируют крахмал и сахарозу по С3-пути.
В С4-пути три фермента, которые регулируются светом, становятся более активными в дневное время суток. Малатдегидрогеназа активируется через тиоредоксин-зависимый восстановительный механизм, показанный на рис. 20-19; ФЕП-карбоксилаза активируется фосфорилированием серинового остатка; а пируват-фосфатдикиназа активируется дефосфорилированием. В последних двух случаях детали инициации светом процессов фосфорилирования или дефосфорилирования неизвестны.
У С4-растений путь ассимиляции СО2 энергетически более дорогой, чем у С3-растений. На каждую молекулу СО2, ассимилированную в С4- пути, должна регенерироваться молекула ФЕП с затратой двух высокоэнергетических фосфатных групп АТР. Таким образом, С4-растениям необходимо пять молекул АТР на ассимиляцию одной молекулы СО2, в то время как С3-растениям надо только три молекулы АТР (девять АТР на один триозофосфат). Однако при увеличении температуры (и уменьшении сродства рубиско к СО2, как было замечено ранее) достигаются условия (около 28-30 °С), при которых выгода устранения фотодыхания значительно превосходит энергетические траты. С4-Растения (например, сорняк росичка кровяная) в течение лета обгоняет в росте большинство С3-растений, что может подтвердить любой опытный садовод.
У CAM-растений фиксация СО2 и активность рубиско разделены во времени
Суккулентные растения, такие как кактусы и ананас, которые в природе произрастают в очень жарком и очень засушливом климате, осуществляют другой вариант фотосинтетической фиксации СО2, уменьшающий потери воды через устьица, через которые СО2 и О2 должны попадать в ткань листа. Вместо пространственного разделения процессов начального захвата СО2 и его фиксации рубиско (как это делают С4-растения) они разобщают эти процессы во времени. Ночью, когда воздух холоднее и более влажный, устьица открыты и пропускают СО2, который затем фиксируется на оксалоацетате ФЕП-карбоксилазой. Оксалоацетат восстанавливается до малата и запасается в вакуолях с целью защиты ферментов цитозоля и пластид от низких значений pH, возникающих из-за диссоциации яблочной кислоты. В дневное время устьица закрыты, что предотвращает потерю воды, которая происходила бы из-за высоких дневных температур, и углерод, включенный ночью в малат, высвобождается NADP-зависимой малатдегидрогеназой в виде СО2. Этот СО2 далее вовлекается ферментом рубиско в цикл Кальвина. Так как этот метод фиксации СО2 впервые был открыт в многолетних толстянковых растениях из семейства Crassulaceae, он был назван crassulacean add metabolism (CAM-метаболизм), а соответствующие растения названы САМ-растениями.
Краткое содержание раздела 20.2 Фотодыхание, С4- и САМ-пути
■ Когда рубиско использует в качестве субстрата О2, а не СО2, образующийся 2-фосфогликолат распадается по кислород-зависимому пути. В результате увеличивается расход О2 — фотодыхание, или, более правильно, цикл фотосинтетического окисления углерода или С2-путь. 2-Фосфогликолат превращается в глиоксилат, затем в глицин, а после этого в серин под действием ферментов стромы хлоропластов, пероксисом и митохондрий.
■ У С4-растений путь ассимиляции углерода сводит к минимуму возможность фотодыхания: в клетках мезофилла СО2 включается в четырехуглеродное соединение, которое попадает в клетки обкладки и высвобождает СО2 в высоких концентрациях. Свободный СО2 фиксируется рубиско и другие реакции цикла Кальвина происходят так же, как и у С3-растений.
■ У CAM-растений в темноте СО2 соединяется с малатом и хранится в вакуолях до наступления дня, когда устьица закрыты (для уменьшения потери влаги), и малат становится источником СО2 для рубиско.