Основы биохимии Том 1 - А. Ленинджер 1985
Биомолекулы
Глобулярные белки: структура и функция гемоглобина
Силы, стабилизирующие третичную структуру глобулярных белков
Мы уже знаем, каким образом формируется и поддерживается вторичная структура полипептидов. Внутрицепочечные водородные связи скрепляют а- спираль, а межцепочечные водородные связи стабилизируют ß-конформацию цепей, образующих складчатые слои (разд. 7.11). При наличии соответствующей аминокислотной последовательности а-спираль и ß-конформация - это самые устойчивые структуры, которые может иметь данный участок цепи; иными словами, они являются структурами с наименьшей свободной энергией. Какие силы стабилизируют третичную структуру глобулярных белков? Существуют четыре типа взаимодействий, сочетание которых обеспечивает правильное взаимное расположение всех витков и петель в глобулярных белках (рис. 8-9) при соблюдении нормальных биологических условий (температура, pH, концентрация ионов).
1. Водородные связи между R-группами остатков, расположенных в соседних петлях полипептидной цепи. Например, гидроксильная группа остатка серина в одном участке полипептидной цепи может образовать водородную связь с атомом азота в кольце остатка гистидина, находящегося в соседней петле той же цепи.
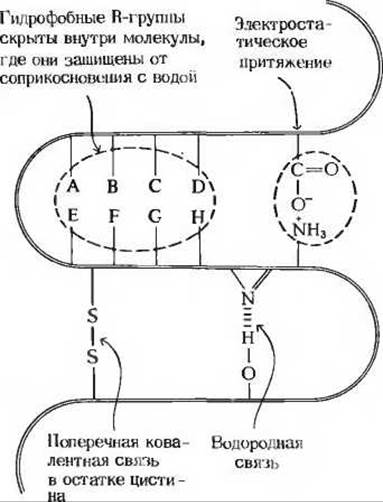
Рис. 8-9. Силы, стабилизирующие третичную структуру глобулярных белков.
2. Электростатическое притяжение между противоположно заряженными R-группами. Например, отрицательно заряженная карбоксильная группа (—СОО—) остатка глутаминовой кислоты может притягиваться положительно заряженной Б-аминогруппой (—NH3) остатка лизина, расположенного в соседней петле.
3. Гидрофобные взаимодействия. Гидрофобные R-группы некоторых аминокислотных остатков (см. табл. 8-1) избегают контактов с водным окружением и стремятся собраться вместе внутри глобулярной структуры, где они защищены от соприкосновения с водой.
4. Ковалентные поперечные связи. Соседние петли полипептидной цепи в некоторых белках, например в рибонуклеазе, содержат остатки цистина, которые образуют внутрицепочечные ковалентные поперечные связи между соседними петлями. Такие ковалентные поперечные связи, конечно, намного прочнее, чем перечисленные выше нековалентные взаимодействия, однако они встречаются не во всех белках. Следовательно, те белки, у которых нет дисульфидных поперечных связей, поддерживают свойственную им третичную структуру при помощи множества слабых нековалентных взаимодействий и разного рода контактов, в совокупности придающих структуре достаточную прочность.
Хотя нативная третичная структура каждого глобулярного белка отвечает минимуму свободной энергии и потому является самой устойчивой конформацией, какую только может принять данная полипептидная цепь, третичную структуру глобулярных белков не следует считать абсолютно жесткой и неподвижной. Многие глобулярные белки в норме претерпевают конформационные изменения при выполнении ими биологических функций. Например, молекула гемоглобина, о котором мы будем говорить дальше, изменяет свою конформацию при связывании кислорода и возвращается к исходной конформации после его освобождения. Кроме того, молекулы многих ферментов претерпевают конформационные изменения при связывании субстратов-это составляет часть их каталитического действия. Полипептидный остов глобулярных белков характеризуется определенной степенью гибкости, вследствие чего эти белки подвержены локальным внутренним флуктуациям, т.е. они как бы «дышат».