Основы биохимии Том 1 - А. Ленинджер 1985
Биомолекулы
Липиды и мембраны
Полярные липиды и белки - основные компоненты мембран
Внешние, или плазматические, мембраны многих клеток, а также мембраны ряда внутриклеточных органелл, например митохондрий и хлоропластов, удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембраны от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах мозга, наоборот, около 80% липидов и только 20% белков. Липидная часть мембран представляет собой смесь различного рода полярных или амфипатических липидов В мембранах животных клеток присутствуют в основном фосфоглицериды и в меньших количествах - сфинголипиды. Триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержат значительные количества холестерола и его эфиров. Для каждого типа мембран любой животной клетки характерен свой относительно постоянный липидный состав (табл. 12-5).
Таблица 12-5. Примерный липидный состав (в процентах) субклеточных мембран печени крысы
Обратите внимание на высокий уровень холестерола и его эфиров, а также гликолипидов (значительную часть которых составляют ганглиозиды) в плазматической мембране.
|
Мембрана |
Фосфолипиды |
Холестерол |
Гликолипиды |
Эфиры холестерола и минорные компоненты |
|
Плазматическая |
57 |
15 |
6 |
22 |
|
Аппарата Гольджи |
57 |
9 |
0 |
34 |
|
Эндоплазматического ретикулума |
85 |
5 |
0 |
10 |
|
Внутренняя митохондриальная |
92 |
0 |
0 |
8 |
|
Ядерная |
85 |
5 |
0 |
10 |
Природные мембраны характеризуются очень малой толщиной (от 6 до 9 нм), эластичностью, а также тем, что они находятся в жидком состоянии. Через мембраны легко проходит вода, но они практически полностью непроницаемы для заряженных ионов типа Na+, Cl или Н+ и для полярных, но не заряженных молекул, например сахаров. Только те полярные молекулы проникают через природные мембраны, для которых существуют специфические транспортные системы, или переносчики. В то же время растворимые в липидах молекулы легко проходят через природные мембраны благодаря своей способности растворяться в углеводородном слое мембран. Как природные мембраны, так и полярные липидные бислои обладают высоким электрическим сопротивлением и потому являются хорошими изоляторами. Такое сходство свойств позволяет считать, что природные мембраны представляют собой сплошной, пластиноподобный полярный липидный бислой, в который включены многочисленные белки.
В различных мембранах на долю белков приходится от 20 до 80% массы. В мембране эритроцита, например, содержится около 20 различных белков, а во внутренней митохондриальной мембране их значительно больше. Некоторые белки в мембранах обладают ферментативной активностью, другие обеспечивают связывание и перенос молекул полярных веществ через мембраны. Мембранные белки различаются по характеру связи с мембранными структурами. Одни белки, называемые внешними, или периферическими, непрочно связаны с поверхностью мембраны; другие, называемые внутренними, или интегральными, - погружены внутрь мембраны и даже могут пронизывать ее насквозь (рис. 12-17). Периферические белки обычно легко экстрагируются из мембран, тогда как интегральные белки могут быть выделены только при помощи детергентов или органических растворителей.
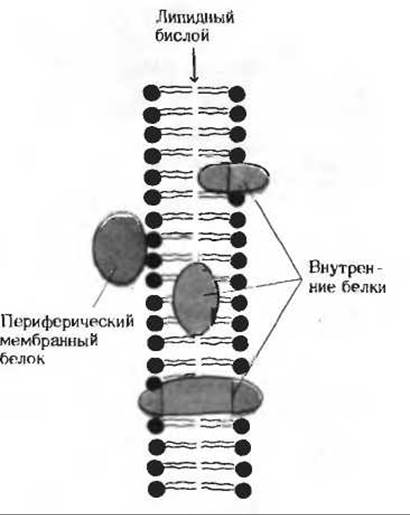
Рис. 12-17. Мембранные белки. Периферические (внешние) белки легко отделяются от мембраны. тогда как интегральные мембранные белки плохо экстрагируются водными растворами.
Важную роль в изучении строения мембран сыграли методы химического анализа, но нараду с этим обширная информация была получена и при использовании электронной микроскопии (дополнение 12-1).
Дополнение 12-1. Электронная микроскопия мембран
В сочетании с разнообразными методами приготовления и окрашивания тканей электронная микроскопия позволила выявить много важных деталей в строении мембран. На приведенных ниже фотографиях видны три разных изображения плазматической мембраны эритроцита, полученные при электронной микроскопии препаратов, приготовленных тремя различными методами.
На рис. 1 показан вид плазматической мембраны эритроцита сбоку. Эта фотография, на которой видны две темные линии («железнодорожные пути») получена после фиксации клеток четырехокисью осмия. Линии соответствуют наружному и внутреннему полярным слоям, состоящим из полярных голов мембранных липидов. Светлая зона между линиями соответствует гидрофобной части липидного бислоя, в которой находятся неполярные углеводородные цепи жирных кислот. Эта микрофотография получена методом трансмиссионной электронной микроскопии.
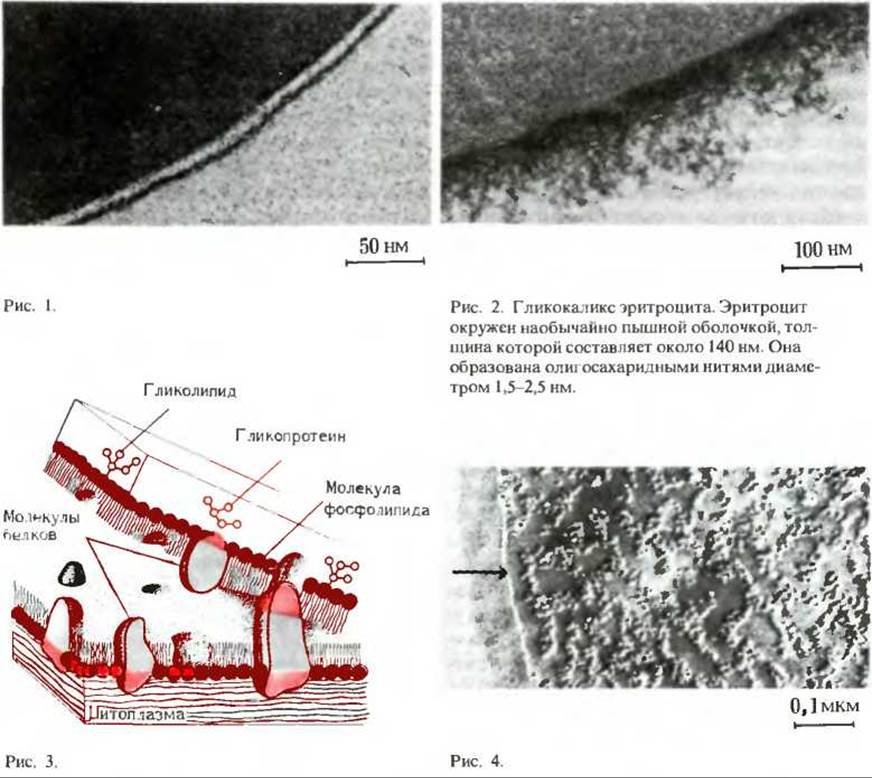
На рис. 2 показан гликокаликс (разд. 11.12) на внешней поверхности эритроцита, выявленный специальным методом окрашивания. Эта «пушистая оболочка» состоит из гидрофильных олигосахаридных групп гликопротеинов и гликолипидов, ее толщина - около 100 нм, что приблизительно в 10 раз превышает толщину липидного бислоя.
На рис. 3 и 4 показана внутренняя сторона мембраны эритроцита; электронная микрофотография на рис. 4 получена с использованием метода замораживания-скалывания. При приготовлении препаратов этим методом клетки сначала замораживают, а затем замороженный блок раскалывают. Иногда линия раскола проходит в плоскости между двумя липидными слоями (рис. 3). С обеих образующихся при этом поверхностей делают отпечатки, которые затем исследуют в электронном микроскопе (рис. 4). Внутренняя часть каждого из липидных слоев имеет гладкую поверхность; расположенные на ней скопления - это молекулы интегральных белков. Стрелкой показан внешний край скола.
При исследовании других особенностей строения мембран применяются также методы сканирующей электронной микроскопии [например, для изучения микроворсинок, расположенных на поверхности клеток (рис. 2-20, разд. 2-19)] и негативного контрастирования [для выявления крупных периферических белков, например, F1-ATP-азы внутренней митохондриальной мембраны (гл. 17)].