Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
Кольцевая ДНК реплицируется в двух направлениях
Мы уже видели, что ДНК бактерий и многих ДНК-содержащих вирусов представляет собой кольцевую двойную спираль. Как только было сделано это открытие, возник вопрос о том, как реплицируется кольцевая ДНК. Расщепляется ли сначала кольцевая ДНК перед репликацией, превращаясь в линейную молекулу, или же она способна реплицироваться в виде кольца? Важные эксперименты, выполненные Джоном Кэрнсом, показали, что ДНК в неповрежденных клетках Е. coli реплицируется, оставаясь в кольцевой форме. Кэрнс выращивал Е. coli на среде, содержащей тимидин, меченный радиоактивным изотопом водорода - тритием (3Н). При этом ДНК в клетках Е. coli становилась радиоактивной. Когда ее осторожно выделяли в релаксированной форме и наносили на фотографическую пластинку, радиоактивные остатки тимидина вызывали на экспонируемой пластинке образование “треков” из зерен серебра, создающих изображение молекулы ДНК. На основании этих изображений Кэрнс заключил, что интактная хромосома - это гигантское кольцо; этот вывод согласуется с полученной ранее генетическими методами кольцевой картой ДНК Е. coli. Однако в радиоактивной ДНК, выделенной из клеток в процессе репликации, была выявлена дополнительная радиоактивная петля (рис. 28-3). Кэрнс предположил, что эта петля в ДНК возникает в результате образования двух радиоактивных дочерних цепей, комплементарных родительским цепям, по мере того как репликативная вилка движется по кольцу родительской ДНК.
Первоначально считали, что репликация начинается в фиксированной точке родительской ДНК (она называется точкой начала репликации) и что единственная репликативная вилка движется по кольцевой молекуле ДНК в одном направлении (рис. 28-3). Однако последующие эксперименты, проведенные на хромосомах Е. coli и вирусов, показали, что репликация обычно происходит в двух направлениях, т. е. существуют две репликативные вилки. Обе вилки возникают в одной точке и удаляются от нее в обоих направлениях одновременно, пока снова не встретятся (рис. 28-3). В этой точке два полностью синтезированных дочерних двухцепочечных кольца разделяются: каждое из них содержит одну старую и одну новую цепь.
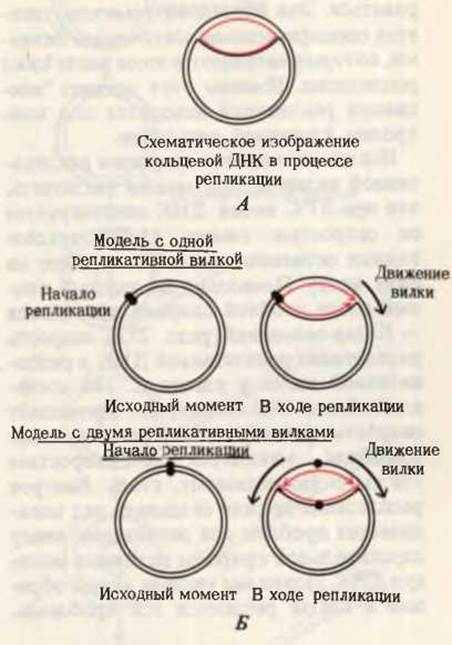
Рис. 28-3. Репликация хромосомы Е. coli. А. Схематическое изображение меченной тритием хромосомы Е. coli в ходе репликации. Б. Интерпретация процесса репликации (новосинтезированные дочерние цепи обозначены красным цветом). Согласно одной модели, от точки начала репликации движется только одна репликативная вилка. Согласно другой модели, в точке начала репликации возникают две репликативные вилки, которые движутся в противоположных направлениях до встречи друг с другом. Хромосомы Е. coli и других бактерий, а также многих ДНК-содержащих вирусов реплицируются в соответствии со второй моделью.
Участок начала репликации представляет собой нуклеотидную последовательность длиной 100-200 пар оснований, без которой ДНК не может реплицироваться. Эта последовательность узнается специфическими клеточными белками, которые начинают в этом месте цикл репликации. Именно этот процесс инициации репликации находится под контролем клеточной регуляции.
Исходя из скорости движения репликативной вилки Е. coli, можно заключить, что при 37°С новая ДНК синтезируется со скоростью свыше 45 000 нуклеотидных остатков в минуту в расчете на одну вилку. Поскольку на каждый полный виток двойной спирали приходится ~ 10 пар оснований (разд. 27.6), скорость расплетания родительской ДНК в репликативной вилке у клеток Е. coli составляет более 4500 об/мин, что превышает скорость вращения вала в двигателе автомобиля, мчащегося со скоростью 110 км/ч. По-видимому, столь быстрое расплетание должно создавать ряд механических проблем для репликации ввиду двуспиральной природы нативных молекул ДНК. Позже мы увидим, каким образом в клетке решаются эти проблемы.