Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Репликация и транскрипция ДНК
У РНК-содержащих вирусов ДНК считывается при помощи обратной транскриптазы
Некоторые онкогенные РНК-содержащие вирусы животных, такие, как вирус саркомы Рауса, имеют уникальный фермент - РНК-зависимую ДНК-полимеразу, часто называемую обратной транскриптазой. После того как такой вирус попадает в клетку-хозяина, этот фермент способен катализировать синтез ДНК, комплементарной по отношению к вирусной РНК, которая играет при этом роль матрицы. В результате образуется ДНК, которая содержит гены, обусловливающие рак; эта ДНК часто встраивается в геном эукариотической клетки-хозяина, где она может в течение многих поколений оставаться в скрытом, т. е. неэкспрессируемом, состоянии (рис. 28-24). При определенных условиях такие бездействующие вирусные гены могут активироваться и вызывать репликацию вируса; при других же условиях они могут способствовать превращению такой клетки в раковую.
Существование обратных транскриптаз в РНК-содержащих онкогенных вирусах было предсказано еще в 1962 г. Говардом Темином из Висконсинского университета, а наличие их в этих вирусах было в конце концов продемонстрировано в 1970 г. как самим Темином, так и независимо от него Дэвидом Балтимором из Массачусетского технологического института. Их открытие привлекло большое внимание главным образом потому, что оно представляло собой доказательство возможности передачи генетической информации в направлении от РНК к ДНК. Оно позволило представить, каким способом включаются в геном клетки-хозяина онкогены, находящиеся в РНК-содержащих вирусах в виде РНК. Именно благодаря этому открытию пришлось по-иному сформулировать центральную догму молекулярной биологии (рис. 28-25). Обладающие обратной транскриптазой РНК-содержащие вирусы называют также ретровирусами (“ретро” по-латыни означает “назад”).
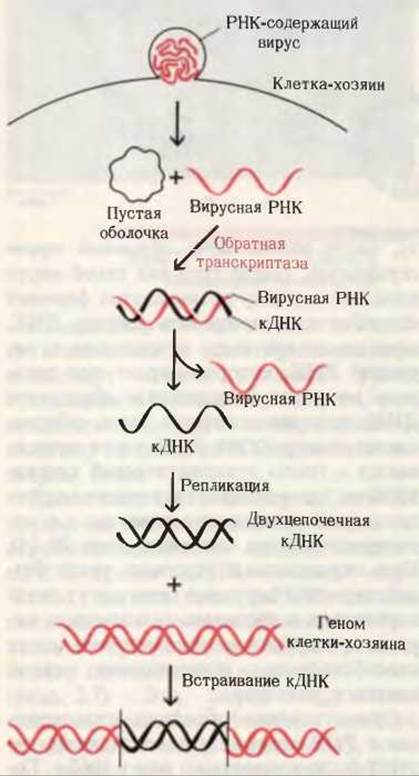
Рис. 28-24. Участие обратной транскриптазы в образовании комплементарной ДНК на вирусной одноцепочечной РНК-матрице в животной клетке. Полученная кДНК может встраиваться в геном клетки-хозяина.
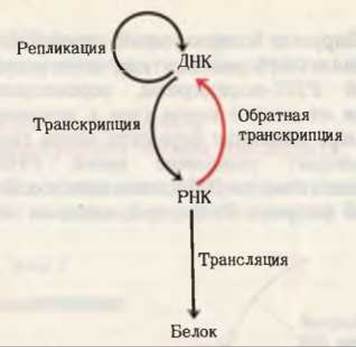
Рис. 28-25. Расширенное толкование центральной догмы молекулярной генетики, учитывающее возможность передачи генетической информации от РНК к ДНК в связи с открытием обратных транскриптаз.
Обнаружение обратной транскриптазы позволило ответить на давний вопрос: как генетическая информация онкогенных РНК-содержащих вирусов может включаться в ДНК клетки-хозяина? Сейчас накапливается все больше данных о том, что в ДНК многих видов животных присутствуют гены, берущие начало от РНК-содержащих вирусов, даже если животные, из которых эта ДНК выделена, сами не подвергались воздействию таких вирусов. Эти наблюдения заставляют предположить, что гены ряда РНК-содержащих вирусов когда-то, возможно на ранних этапах биологической эволюции этих видов, транскрибировались в ДНК и встроились в хромосомы предков этих животных. После этого они передавались из поколения в поколение при репликации всей ДНК клетки, в том числе и содержащихся в ней онкогенов, исходно принадлежавших вирусной РНК. Действительно, согласно одной из теорий происхождения рака, считается, что все мы несем в хромосомах “спящие”, неэкспрессируемые, онкогены, которые попали в геном наших предков в виде РНК-содержащих вирусов, возможно, тысячи или миллионы лет назад. Эта теория далее предполагает, что такие онкогены обычно не транскрибируются, но если их активировать, например, воздействием канцерогенных агентов, то они транскрибируются и транслируются с образованием продуктов, вызывающих трансформацию нормальных клеток человека в злокачественные.
Вирусные обратные транскриптазы, так же как все ДНК- и РНК-полимеразы, содержат ионы Zn2+. Они проявляют наибольшую активность при использовании в качестве матрицы РНК своего вируса, но способны синтезировать также ДНК, комплементарную к самым разным РНК. Обратным транскриптазам необходима затравка; синтез новой цепи ДНК они ведут в направлении 5'→3' и вообще во многих отношениях напоминают ДНК-полимеразы.
Обратная транскриптаза, подобно рестриктирующим эндонуклеазам (разд. 27.24), стала, как мы увидим дальше, очень важным биохимическим инструментом при изучении взаимоотношений ДНК-РНК, а также процесса клонирования ДНК. Этот фермент сделал возможным искусственный синтез ДНК, комплементарной любой матрице РНК, будь то мРНК, тРНК или рРНК. Синтетическую ДНК, полученную таким путем, называют комплементарной ДНК (кДНК). С помощью обратной транскриптазы можно, например, получить синтетический ген (т. е. кДНК), кодирующий одну из полипептидных цепей гемоглобина, исходя из ее мРНК. мРНК, кодирующие цепи гемоглобина, легко могут быть выделены из эритроцитов. В этом и во многих других случаях, когда выделение природного гена, кодирующего какой-либо полипептид эукариот, довольно сложно, а его мРНК доступна, можно получить синтетический ген этой мРНК при помощи обратной транскриптазы. Позже мы увидим, как кДНК используются для клонирования рекомбинантных ДНК (гл. 30).