Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Синтез белка и его регуляция
Инициирующей аминокислотой у прокариот служит N-формилметионин, а у эукариот-метионин
У Е. coli и у всех других прокариот начальным, N-концевым аминокислотным остатком всегда оказывается остаток N-формилметионина (рис. 29-10). В процесс биосинтеза белка он вступает в составе N-формилметионил-тРНК (обозначаемой как фМет-тРНКfMet), которая образуется в результате двух последовательных реакций. Сначала метионин присоединяется с помощью метионил-тРНК-синтетазы к особой инициирующей метиониновой тРНК-тРНКfMet:
![]()
Во второй реакции формильная группа при помощи специфической трансформилазы переносится от донора N-формилтетрагидрофолата (разд. 10.10) к аминогруппе метионинового остатка:

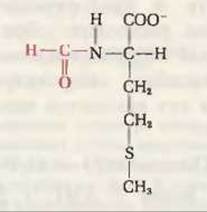
Рис. 29-10. N-формилметионин-инициирующая аминокислота у всех прокариот. N-формильная группа показана красным цветом.
Трансформилаза не способна формилировать свободный метионин. Существуют два вида тРНК, специфичных к метионину,-тРНКMet и тРНКfМеt. Обе эти тРНК могут присоединять метионин в реакции активации, но приобретать формильную группу и становиться инициирующей аминокислотой метионин способен только в составе метионил-тРНКfMet. Другая тРНК-метионил-тРНК используется для встраивания метионина во внутренние участки полипептидной цепи. Блокирование аминогруппы метионина N-формильным остатком препятствует включению такой аминокислоты во внутренние участки цепи, но в то же время позволяет фМет-тРНКfМеt связываться с особым местом инициации на рибосоме, с которым не может связываться ни Мет-тРНКМеt, ни любая другая аминоацил-тРНК.
Что касается эукариотических клеток, то все полипептиды, синтезируемые их внемитохондриальными рибосомами, начинаются с остатка метионина, который поступает туда в составе специальной инициирующей метионил-тРНК. Полипептиды же, синтезируемые в митохондриях и хлоропластах эукариотических клеток так же, как и в бактериях, начинаются с N-формилметионина. Это и другие похожие свойства белоксинтезирующих аппаратов митохондрий и хлоропластов, с одной стороны, и бактерий - с другой, служат подтверждением точки зрения (разд. 2.8 и 17.18), согласно которой митохондрии и хлоропласта произошли от бактерий на ранних этапах эволюции эукариотических клеток. Следует, однако, отметить, что в некоторых других отношениях между механизмами транскрипции и трансляции у бактерий и митохондрий имеются значительные различия.
У нас осталась без ответа еще одна загадка. Поскольку для метионина известен только один кодон, а именно (5')AUG(3'), возникает вопрос: каким образом этот единственный кодон используется для встраивания и начального N-формилметионинового остатка (или метионинового, в случае эукариот), и тех метиониновых остатков, которые предназначены для включения во внутренние участки полипептидных цепей? Ответ на этот вопрос будет дан позже, после рассмотрения этапа инициации белкового синтеза, а пока необходимо остановиться на структуре рибосом.