Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Синтез белка и его регуляция
Инициация синтеза полипептида происходит в несколько стадий
Для инициации полипептидной цепи в клетках прокариот необходимы: 1) 30S-субчастица, содержащая 16S-pPHK; 2) мРНК, кодирующая синтезируемый полипептид; 3) инициирующая N-формилметионил-тРНКfMеt; 4) три белка, называемые факторами инициации (IF-1, IF-2 и IF-3); 5) GTP (табл. 29-1).
Образование инициирующего комплекса протекает в три стадии. На первой из них 30S-рибосомная субчастица связывает фактор инициации 3 (IF-3), который препятствует объединению 30S- и 50S-субчастиц. Затем к 30S-субчастице присоединяется мРНК таким образом, что инициирующий кодон мРНК (5') AUG(3') связывается с определенным участком 30S-субчастицы (рис. 29-14). Правильное расположение инициирующего кодона AUG на 30S-субчастице обеспечивается с помощью особого инициирующего сигнала, представляющего собой участок мРНК, расположенный с 5'-стороны от кодона AUG. Этот сигнал состоит преимущественно из остатков А и G и включает обычно от 6 до 8 таких остатков. Он узнается комплементарной последовательностью 16S-pPHK 30S-субчастицы и благодаря этому мРНК фиксируется в нужном для инициации трансляции положении. Поскольку и для инициирующих, и для внутренних остатков метионина существует всего лишь один кодон, инициирующий сигнал с 5'-стороны от AUG указывает место, с которым надлежит связаться фМет-тРНКfMet. Внутренние кодоны AUG специфичны по отношению к Мет-тРНКMet и не способны связывать фМет-тРНКfMet.
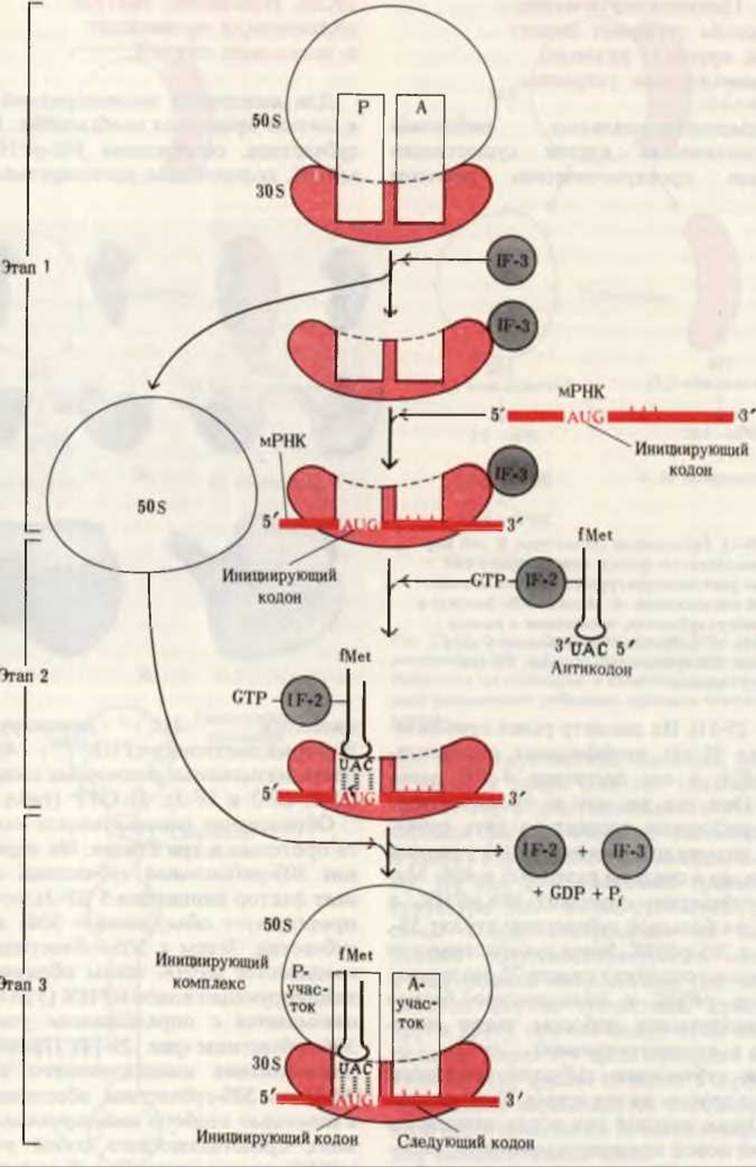
Рис. 29-14. Три стадии процесса образования инициирующего комплекса, протекающего за счет энергии гидролиза GTP до GDP и Рi. IF-1, IF-2 и IF-3-факторы инициации. Буквами Р и А обозначают соответственно пептидильный и аминоацильный участки рибосомы. Роль IF-1 до конца не выяснена.
На второй стадии процесса инициации (рис. 29-14) размер комплекса, состоящего из 30S-субчастицы, IF-3 и мРНК, увеличивается в результате соединения с фактором инициации IF-2, уже связанного с GTP и с инициирующей N-формилметионил-тРНКfMet, которая попадает точно на инициирующий кодон.
На третьей стадии инициации этот большой комплекс взаимодействует с 50S-рибосомной субчастицей; одновременно молекула GTP, связанная с IF-2, гидролизуется до GDP и фосфата, которые высвобождаются из комплекса. Факторы инициации IF-3 и IF-2 также покидают рибосому. Теперь мы имеем функционально активную 70S-рибосому, которая называется инициирующим комплексом; она содержит мРНК и инициирующую N-формилметионил-тРНКfMet. Правильное положение N-формилметионил - тРНКfMet в полном 70S- инициирующем комплексе обеспечивается двумя точками узнавания и связывания. Во-первых, антикодоновый триплет инициирующей аминоацил-тРНК образует комплементарные пары с антипараллельно расположенным кодоновым триплетом AUG в мРНК. Во-вторых, инициирующая аминоацил-тРНК присоединяется к пептидильному P-участку рибосомы. В рибосоме имеется два участка связывания аминоацил-тРНК. аминоацил-, или A-участок и пептидил-, или P-участок. Оба они образованы благодаря специфическому сочетанию областей 30S- и 50S-субчастиц. Инициирующая фМет-тРНК может связываться только с P-участком (рис. 29-14), однако это исключение: все остальные вновь поступающие аминоацил-тРНК присоединяются к A-участку, тогда как Р-участок - это такое место рибосомы, с которого уходят “пустые” (т. е. освободившиеся от аминокислот) тРНК и к которому оказывается прикрепленной растущая пептидил-тРНК.
Инициирующий комплекс теперь готов к процессу элонгации.