Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
Еще о генах: репарация, мутации, рекомбинация и клонирование
Гены из разных организмов можно искусственным образом объединить
Мы рассмотрели несколько вариантов процесса генетической рекомбинации, реализующихся в природных условиях в клетках различных типов. Рекомбинацию генов или наборов генов можно провести также и в пробирке и при этом получить новые комбинации генов, не существующие в природе. Например, из двух разных видов организмов можно выделить гены, кодирующие два различных белка, соединить их вместе и получить новое сочетание генов. Такие искусственные рекомбинантные ДНК служат исключительно ценным инструментом в генетических исследованиях. Кроме того, как мы увидим позже, они могут использоваться и в практических целях.
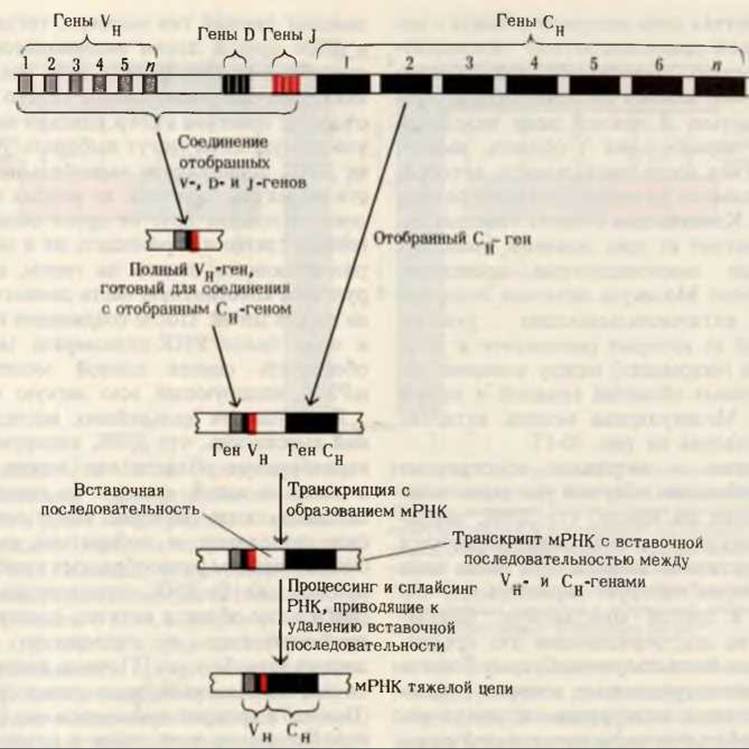
Рис. 30-18. Схема показывает, каким образом ген тяжелой цепи антитела может быть собран в результате перемещения генов V, D и J из разных участков генома; в конце концов образуется полный VH-ген, соединенный С СH-геном. Затем транскрипт мРНК подвергается процессингу, в ходе которого удаляются спейсерные участки и образуется зрелая мРНК, кодирующая тяжелую цепь.
Развитие методов выделения генов и соединения их в новых сочетаниях стало главным биохимическим достижением, открывшим новую эру в генетических исследованиях.
Обнаружение рестриктирующих эндонуклеаз стало первым шагом на пути решения проблемы искусственной рекомбинации генов. Предположим, что мы хотим соединить вместе две двухцепочечные ДНК, выделенные из разных организмов (рис. 30-19). Каждую из них в отдельности обрабатывают одной и той же рестриктирующей эндонуклеазой, которая разрывает обе цепи ДНК с образованием в месте разрыва выступающих концов (разд. 27.24). Допустим, что в каждой из этих ДНК есть лишь один участок, узнаваемый данной рестриктазой. Следовательно, нуклеотидные последовательности выступающих концов двух расщепленных ДНК будут комплементарны. Если теперь смешать эти ДНК, нагреть и медленно охладить, то их липкие концы образуют комплементарные пары оснований, в результате чего возникнет новая рекомбинантная ДНК, цепи которой имеют единичные разрывы (рис. 30-19). При обработке таких ДНК ДНК-лигазой в присутствии источников энергии образуется новая ковалентно сшитая рекомбинантная ДНК.
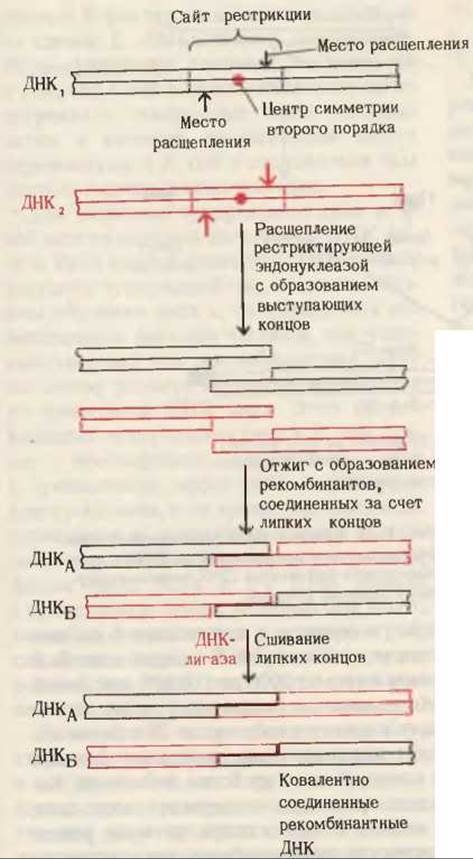
Рис. 30-19. Использование специфической рестриктирующей эндонуклеазы, которая образует в месте разрыва выступающие концы, для расщепления двух разных ДНК (ДНК, и ДНК,) и получения фрагментов для последующей рекомбинации. Поскольку многие рестриктирующие эндонуклеазы расщепляют лишь специфические участки ДНК, обладающие осевой симметрией второго порядка, образующиеся липкие концы комплементарным образом взаимодействуют с концами любой другой ДНК, разрезанной той же самой эндонуклеазой. При отжиге из соответствующих фрагментов ДНК, и ДНК, могут снова образоваться исходные молекулы или же новые рекомбинантные ДНК, как это показано на рисунке. Ковалентное сшивание фрагментов осуществляется под действием ДНК-лигазы.
Другой ключевой фермент, широко применяемый для соединения фрагментов ДНК, - это терминальная трансфераза, которая способна присоединять к 3'-концу цепей ДНК большое число следующих друг за другом дезоксирибонуклеотидных остатков. Этот фермент неспецифичен и может использовать в качестве предшественников dATP, dTTP, dGTP или dCTP. Поскольку для действия терминальной трансферазы не нужна матрица, она способна построить 3'-концевые последовательности, состоящие из нуклеотидов одного типа. Следовательно, к 3'-концам одной из двухцепочечной ДНК можно добавить poly (G)-хвосты, а к 3'-концам другой - poly (С)-хвосты. Поскольку такие хвосты комплементарны друг другу, с их помощью можно соединить две ДНК за счет образования комплементарных пар между основаниями их липких концов (рис. 30-20), ковалентная сшивка соединенных таким образом ДНК осуществляется ДНК-лигазой.
С помощью этих и других ферментов уже соединены многие ДНК из самых разных источников. Одним из первых достижений в этом направлении было встраивание гена рРНК, выделенного из шпорцевой лягушки Xenopus laevis, в плазмиду Е. coli. В другом раннем эксперименте ДНК обезьяньего вируса SV40 (разд. 27.29) была встроена в ДНК фага λ, т. е. были объединены хромосомы животного и бактериального вирусов. С тех пор в лабораторных условиях были получены сотни различных искусственных рекомбинантных ДНК.