Основы биохимии Том 3 - А. Ленинджер 1985
Некоторые аспекты биохимии человека
Пищеварение, транспорт питательных веществ и взаимосвязь обменных процессов
Пища подвергается ферментативному перевариванию, что подготавливает ее к последующему всасыванию
Эта глава первая из трех, освещающих биохимические аспекты метаболизма и его регуляции у человека. Мы начнем с анализа биохимии пищеварения и всасывания продуктов питания в пищеварительном тракте, распределения питательных веществ между различными органами и взаимосвязей обмена веществ в разных тканях. Далее мы перейдем к биохимическим механизмам, посредством которых доставляется в ткани кислород, а также выводятся двуокись углерода и другие конечные продукты метаболизма.
Одним из наиболее важных практических применений биохимии является диагностика заболеваний, сопровождающихся нарушением обмена веществ. На примере такой болезни обмена веществ, как сахарный диабет, мы увидим, какую важную роль играют биохимические анализы в медицине.
В процессе пищеварения в желудочно-кишечном тракте млекопитающих три основных компонента пищи - углеводы, жиры и белки - подвергаются ферментативному гидролизу, распадаясь при этом на составляющие строительные блоки, из которых они образованы. Этот процесс необходим для утилизации пищевых продуктов, поскольку клетки, выстилающие кишечник, способны всасывать в кровоток только относительно небольшие молекулы. Например, усвоение полисахаридов и даже дисахаридов становится возможным только после их полного гидролиза пищеварительными ферментами до моносахаридов. Аналогичным образом белки и липиды также должны быть гидролизованы до блоков, из которых они построены.
На рис. 24-1 приведена схема пищеварительной системы человека. Процесс пищеварения начинается с ротовой полости и желудка, тогда как конечные этапы переваривания всех основных компонентов пищи и всасывание в кровь составляющих их структурных блоков происходят в тонком кишечнике. Анатомически тонкий кишечник хорошо приспособлен для выполнения этой функции, поскольку он обладает очень большой площадью поверхности, через которую происходит всасывание. Тонкий кишечник характеризуется не только большой длиной (4-4,5 м), но также наличием на его внутренней поверхности множества складок с большим количеством пальцевидных выступов, называемых ворсинками. Каждая ворсинка покрыта эпителиальными клетками, несущими многочисленные микроворсинки (рис. 24-2). Ворсинки создают огромную поверхность, через которую продукты переваривания быстро транспортируются в эпителиальные клетки, а из них - в капилляры кровеносной системы и в лимфатические сосуды, расположенные в стенке кишечника. Площадь поверхности тонкого кишечника человека составляет ~180 м2, т. е. лишь немногим меньше игровой площадки теннисного корта.
В микроворсинках содержатся пучки актиновых микрофиламентов (разд. 7.19), соединенных в основаниях микроворсинок с сетью миозиновых нитей. Эта система нитей обеспечивает волнообразные колебания микроворсинок, благодаря которым происходит местное перемешивание и лучшее всасывание переваренных питательных веществ.
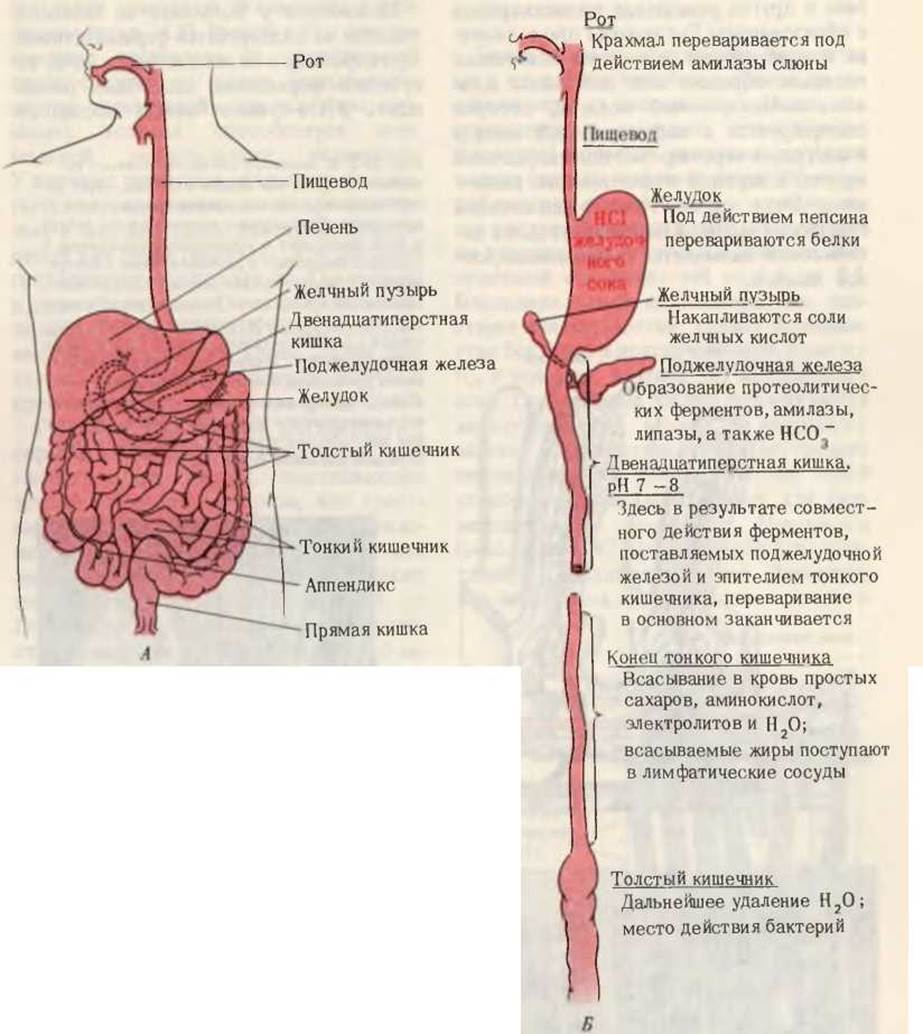
Рис. 24-1. А. Желудочно-кишечный тракт человека. Б. Последовательные этапы переваривания и всасывания.
а. Переваривание углеводов
У человека из углеводов перевариваются в основном полисахариды - крахмал и целлюлоза, содержащиеся в растительной пище, и гликоген, содержащийся в пище животного происхождения. Крахмал и гликоген полностью расщепляются ферментами желудочно-кишечного тракта до составляющих их структурных блоков, а именно свободной D-глюкозы. Этот процесс начинается во рту во время пережевывания пищи благодаря действию фермента амилазы, выделяемого слюнными железами. Амилаза слюны гидролизует многие из а(1→4)-гликозидных связей в крахмале и в гликогене. При этом образуется смесь, состоящая из мальтозы, глюкозы и олигосахаридов. Когда мы жуем сухари или крекеры, они становятся постепенно слаще, поскольку содержащийся в них безвкусный крахмал подвергается ферментативному гидролизу с образованием сахаров. Переваривание крахмала, гликогена и других усвояемых полисахаридов с образованием D-глюкозы продолжается и завершается в тонком кишечнике главным образом под действием амилазы поджелудочной железы, которая синтезируется в поджелудочной железе и поступает через проток поджелудочной железы в верхний отдел тонкого кишечника. Этот отдел тонкого кишечника с наиболее высокой пищеварительной активностью называется двенадцатиперстной кишкой.
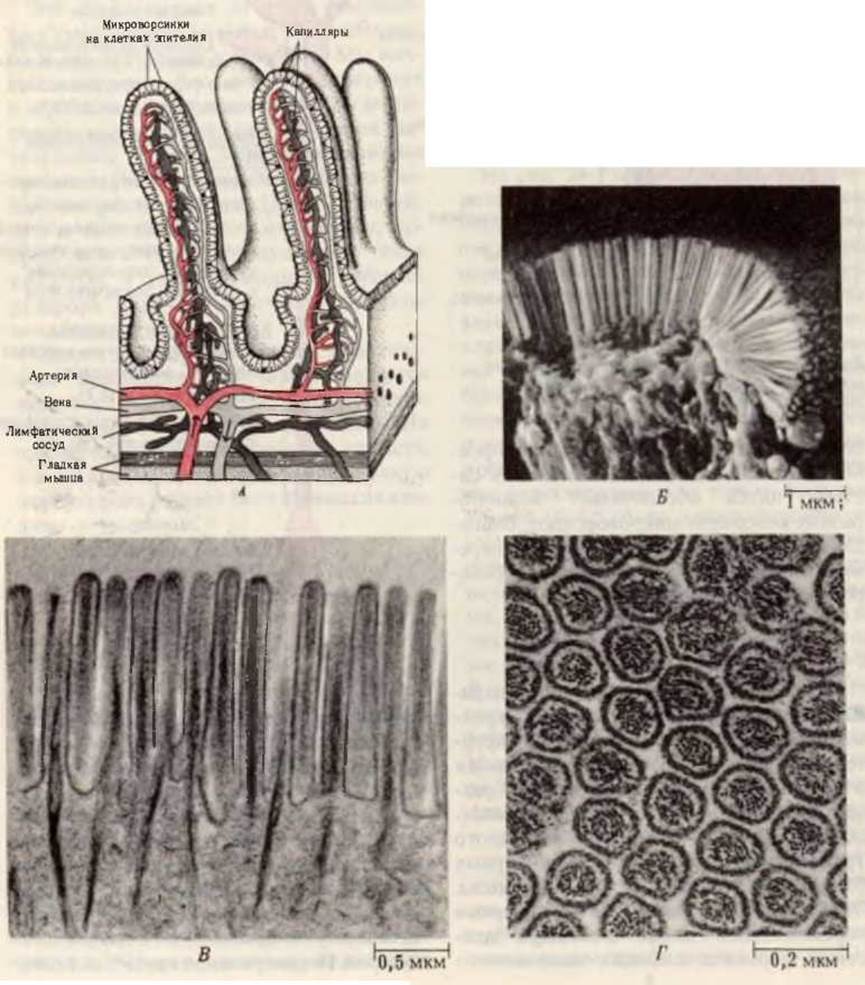
Рис. 24-2. А. Ворсинки слизистой тонкого кишечника; видно, как велика площадь, через которую происходит всасывание продуктов пищеварения. Всасываемые аминокислоты, сахара и соли поступают в кровеносные капилляры, а триацилглицеролы - в расположенные в центре ворсинок лимфатические сосуды. Каждая эпителиальная клетка несет большое число микроворсинок. Б. Микрофотография ворсинок, полученная с помощью сканирующего электронного микроскопа. В, Г. Микрофотографии соответственно продольного и поперечного срезов ворсинок, полученные с помощью трансмиссионного электронного микроскопа; видны внутренние микрофиламенты, обеспечивающие волнообразное движение ворсинок.
Целлюлоза у большинства млекопитающих не подвергается ферментативному гидролизу и не используется из-за отсутствия ферментов, способных расщеплять ß(1→4)-связи между последовательными остатками D-глюкозы в целлюлозе (разд. 11.8). Вместе с тем непереваренная целлюлоза из растительной пищи создает ту массу (называемую иногда “клетчаткой” или “грубым кормом”), которая способствует нормальной перистальтике кишечника. У жвачных животных целлюлоза подвергается перевариванию, но не прямым путем, а под действием бактерий, находящихся в их рубце (желудке). Эти бактерии гидролизуют целлюлозу до D-глюкозы и далее сбраживают D-глюкозу до лактата, ацетата и пропионата, которые всасываются и поступают в кровь. Далее лактат и пропионат в печени жвачных превращаются в сахар крови (разд. 20.10).
Гидролиз дисахаридов катализируют ферменты, находящиеся в наружном крае эпителиальных клеток, выстилающих тонкий кишечник. Сахароза, или тростниковый сахар, гидролизуется с образованием D-глюкозы и D-фруктозы под действием сахаразы, называемой также инвертазой; лактоза гидролизуется до D-глюкозы и D-галактозы под действием лактазы, называемой также ß-галактозидазой; в результате гидролиза мальтозы под действием мальтазы образуются две молекулы D-глюкозы. Напомним (разд. 11.5), что многим представителям азиатских и африканских рас во взрослом состоянии свойственна непереносимость лактозы, обусловленная исчезновением в их тонком кишечнике лактазной активности, имевшейся в грудном и детском возрасте. У людей с непереносимостью лактозы этот сахар остается в кишечнике в нерасщепленном виде и часть его подвергается сбраживанию под действием микроорганизмов. Это вызывает диаррею и образование газов в кишечнике.
В эпителиальных клетках, выстилающих тонкий кишечник, D-фруктоза, D-галактоза и D-манноза частично превращаются в D-глюкозу (разд. 15.9). Смесь всех этих простых гексоз поглощается эпителиальными клетками, выстилающими тонкий кишечник, и доставляется кровью в печень.
б. Переваривание белков
Белки пищи расщепляются ферментами в желудочно-кишечном тракте до составляющих их аминокислот. Белки, поступающие в желудок, стимулируют выделение гормона гастрина, который в свою очередь вызывает секрецию соляной кислоты обкладочными клетками желез слизистой желудка (рис. 24-3), а также пепсиногена главными клетками. Желудочный сок имеет pH от 1,5 до 2,5. Благодаря такой кислотности он действует как антисептик, убивая большинство бактерий и других клеток. Кроме того, в условиях низкого pH желудочного сока глобулярные белки подвергаются денатурации, их молекулы разворачиваются и вследствие этого внутренние пептидные связи полипептидных цепей становятся более доступными для ферментативного гидролиза. Пепсиноген (мол. масса 40000), являющийся неактивным предшественником фермента, или зимогеном, превращается в желудочном соке в активный пепсин в результате ферментативного действия самого пепсина, т.е. путем автокатализа. В ходе этого процесса (рис. 24-4) с N-конца полипептидной цепи пепсиногена отщепляются 42 аминокислотных остатка в виде смеси коротких пептидов. Остающаяся интактной остальная часть молекулы пепсиногена представляет собой ферментативно активный пепсин (мол. масса 33 000). В желудке пепсин гидролизует те пептидные связи в белках, которые образованы ароматическими аминокислотами - тирозином, фенилаланином и триптофаном, а также рядом других (табл. 24-1); в итоге из длинных полипептидных цепей образуется смесь более коротких пептидов.
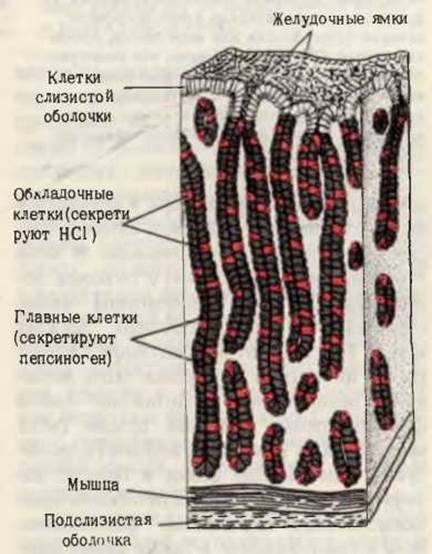
Рис. 24-3. Железы слизистой желудка. Обкладонные клетки секретируют HCl под действием гормона гастрина, который продуцируется эпителиальными клетками при поступлении в желудок белков. Главные клетки секретируют пепсиноген.
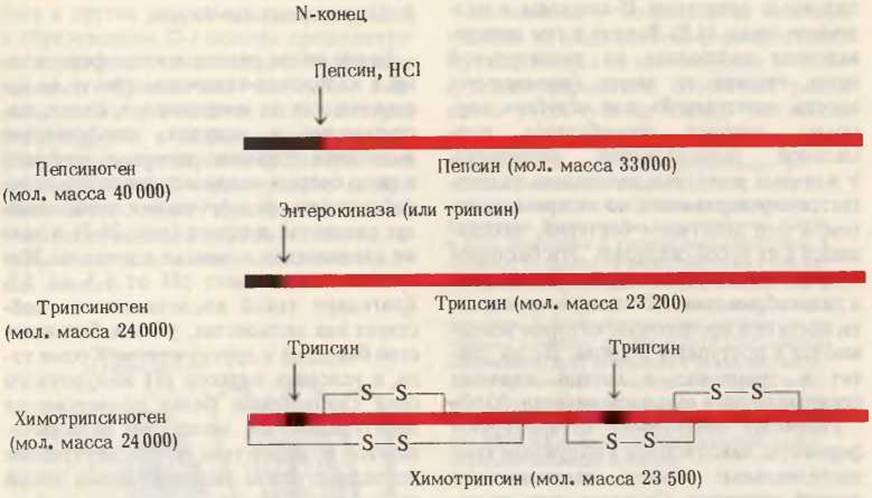
Рис. 24-4. Активация зимогенов пепсина, трипсина и химотрипсина. На диаграммах показаны участки зимогенов, подвергающиеся протеолизу, в результате которого высвобождаются активные ферменты (показаны красным). Те фрагменты полипептидных цепей зимогенов, которые отщепляются или вырезаются, показаны черным. Обратите внимание, что химотрипсин состоит из трех полипептидных цепей, ковалентно связанных друг с другом двумя дисульфидными связями и нековалентно - за счет водородных связей и гидрофобных взаимодействий (дополнение 9-4).
Как только кислое содержимое желудка попадает в тонкий кишечник, в нем под влиянием низкого pH начинается секреция гормона секретина, поступающего в кровь. Этот гормон в свою очередь стимулирует выделение из поджелудочной железы в тонкий кишечник бикарбоната, что приводит к нейтрализации НСl желудочного сока. В результате pH резко возрастает от 1,5-2,5, до ~ 7. В тонком кишечнике переваривание белков продолжается. Поступление аминокислот в двенадцатиперстную кишку вызывает освобождение гормона холецистокинина (разд. 25.22), который стимулирует секрецию нескольких ферментов поджелудочной железы с оптимумом pH около 7. Три из них - трипсин, химотрипсин и карбоксипептидаза - вырабатываются экзокринными клетками поджелудочной железы (рис. 24-5) в виде ферментативно неактивных зимогенов — соответственно трипсиногена, химотрипсиногена и прокарбоксипептидазы. Благодаря синтезу протеолитических ферментов в виде неактивных предшественников экзокринные клетки не подвергаются разрушению этими ферментами. Попав в тонкий кишечник, трипсиноген превращается в активную форму-трипсин (рис. 24-4) - под действием энтерокиназы, специализированного протеолитического фермента, секретируемого клетками кишечного эпителия. Свободный трипсин по мере своего образования также участвует в каталитическом превращении трипсиногена в трипсин. Образование свободного трипсина обусловлено отщеплением гексапептида от N-конца полипептидной цепи трипсиногена. Как мы уже видели (разд. 6.7), трипсин гидролизует пептидные связи, образованные с участием карбонильных групп лизина и аргинина (табл. 24-1).
Таблица 24-1. Ферменты, участвующие в переваривании белков, и их специфичность в отношении пептидных связей, образуемых разными аминокислотными остатками
Пепсин |
Туr, Phe, Тrр, а также Leu, Glu, Gln |
Трипсин |
Lys, Arg |
Химотрипсин |
Tyr, Phe, Trp |
Карбоксипептидаза |
|
Последовательное отщепление С-концевых остатков |
|
Аминопептидаза |
|
Последовательное отщепление N-концевых остатков (за исключением остатков пролина) |
Молекула химотрипсиногена представляет собой одну полипептидную цепь с несколькими внутрицепочечными дисульфидными связями. Попав в тонкий кишечник, химотрипсиноген превращается в химотрипсин под действием трипсина, который разрывает длинную полипептидную цепь химотрипсиногена в двух местах, выстригая дипептиды (рис. 24-4). Три фрагмента, образовавшиеся из исходной цепи химотрипсиногена, удерживаются, однако, вместе посредством перекрестных дисульфидных связей (дополнение 9-4). Химотрипсин гидролизует пептидные связи, образованные остатками фенилаланина, тирозина и триптофана (табл. 24-1). Следовательно, трипсин и химотрипсин расщепляют полипептиды, образовавшиеся в желудке под действием пепсина, на пептиды меньшей величины. Этот этап переваривания белков протекает с очень высокой эффективностью, поскольку пепсин, трипсин и химотрипсин проявляют при гидролизе полипептидных цепей разную специфичность в отношении пептидных связей, образованных разными аминокислотами.
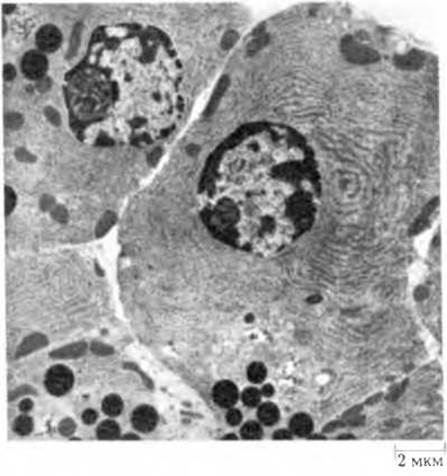
Рис. 24-5. Экзокринные клетки поджелудочной железы. Цитоплазма клеток целиком заполнена шероховатым эндоплазматическим ретикулумом. Находящиеся на его мембранах рибосомы синтезируют полипептидные цепи зимогенов многих пищеварительных ферментов. Зимогены накапливаются в вакуолях, превращающихся в конце концов в зрелые зимогенные гранулы. При стимуляции клетки ее плазматическая мембрана сливается с мембраной, окружающей зимогенные гранулы; последние видны в нижней части рисунка в виде темных частиц сферической формы. Содержимое гранул высвобождается в просвет протока (светлая область в нижней левой части рисунка) посредством зкзоцитоза. Отдельные протоки в конечном итоге ведут в общий проток поджелудочной железы и далее в тонкую кишку.
Деградация коротких пептидов в тонком кишечнике осуществляется другими пептидазами. К ним относится в первую очередь карбоксипептидаза - цинк содержащий фермент (разд. 10.21), синтезируемый в поджелудочной железе в виде неактивного зимогена прокарбоксипептидазы. Карбоксипептидаза последовательно отщепляет от пептидов С-концевые остатки. Тонкий кишечник секретирует также аминопептидазу, отщепляющую от коротких пептидов один за другим N-концевые остатки (табл. 24-1). В результате последовательного действия этих протеолитических ферментов и пептидаз перевариваемые белки в конечном итоге превращаются в смесь свободных аминокислот, которые далее транспортируются через эпителиальные клетки, выстилающие тонкие кишки. Свободные аминокислоты проникают в капилляры ворсинок и переносятся кровью в печень.
В желудочно-кишечном тракте человека не все белки перевариваются целиком (гл. 26). Большинство животных белков почти полностью гидролизуются до аминокислот, однако ряд фибриллярных белков, например кератин, переваривается только частично. Многие белки растительной пищи, в частности белки зерен злаков, неполностью расщепляются в силу того, что белковая часть семян и зерен покрыта неперевариваемой целлюлозной оболочкой (шелухой).
Известно редкое заболевание стеаторрея (упорный понос), при котором ферменты кишечника не способны переваривать определенные водорастворимые белки зерна, в частности глиадин, повреждающий эпителиальные клетки кишечника. Понятно, что из пищи таких больных следует исключить зерновые продукты. Другим заболеванием, связанным с отклонением от нормы активности протеолитических ферментов пищеварительного тракта, является острый панкреатит. При этом заболевании, обусловленном нарушением процесса выделения сока поджелудочной железы в кишечник, предшественники протеолитических ферментов (зимогены) превращаются в соответствующие каталитически активные формы слишком рано, будучи еще внутри клеток поджелудочной железы.
В результате эти мощные ферменты воздействуют на ткань самой железы, вызывая глубокое и очень болезненное разрушение органа, что может привести к смертельному исходу. В норме зимогены, выделяемые поджелудочной железой, не активируются до тех пор, пока не попадут в тонкий кишечник. Поджелудочная железа защищается от самопереваривания и другим путем: в ней синтезируется особый белок - специфический ингибитор трипсина. Поскольку свободный трипсин активирует не только трипсиноген и химотрипсиноген, но также и зимогены двух других пищеварительных ферментов - прокарбоксипептидазу и проэластазу, - ингибитор трипсина успешно предотвращает преждевременное образование свободных протеолитических ферментов в клетках поджелудочной железы.
в. Переваривание жиров
Переваривание триацилглицеролов (нейтральных жиров) начинается в тонком кишечнике, куда из поджелудочной железы поступает зимоген пролипаза. Здесь пролипаза превращается в активную липазу, которая в присутствии желчных кислот (см. ниже) и специального белка, называемого колипазой, присоединяется к капелькам триацилглицеролов и катализирует гидролитическое отщепление одного или обоих крайних жирнокислотных остатков с образованием смеси свободных жирных кислот в виде их Na+ - или К+ -солей (мыл) и 2-моноацилглицеролов (рис. 24-6). Небольшое количество триацилглицеролов остается при этом негидролизованным.
Образовавшиеся мыла и нерасщепленные ацилглицеролы эмульгируются в виде мелких капелек под действием перистальтики (перемешивающие движения кишечника), а также под влиянием солей желчных кислот и моноацилглицеролов, которые являются амфипатическими соединениями и потому функционируют как детергенты. Жирные кислоты и моноацилглицеролы из этих капелек поглощаются кишечными клетками, где из них в основном вновь синтезируются триацилглицеролы (разд. 21.8). Далее триацилглицеролы проникают не в капилляры крови, а в небольшие лимфатические сосуды кишечных ворсинок - лактеали (иначе - млечные, или хилёзные, сосуды). Оттекающая от тонких кишок лимфа, называемая хилус (млечный сок), после переваривания жирной пищи напоминает по виду молоко из-за обилия взвешенных в ней хиломикронов - мельчайших капелек эмульгированных триацилглицеролов диаметром около 1 мкм. Хиломикроны имеют гидрофильную оболочку, состоящую из фосфолипидов и специального белка, который удерживает хиломикроны во взвешенном состоянии. Хиломикроны проходят через грудной проток в подключичную вену (рис. 24-7). После потребления жирной пищи даже плазма крови становится опалесцирующей из-за высокой концентрации в ней хиломикронов, но эта опалесценция исчезает через 1-2 ч, так как триацилглицеролы выводятся из крови, поступая главным образом в жировую ткань.
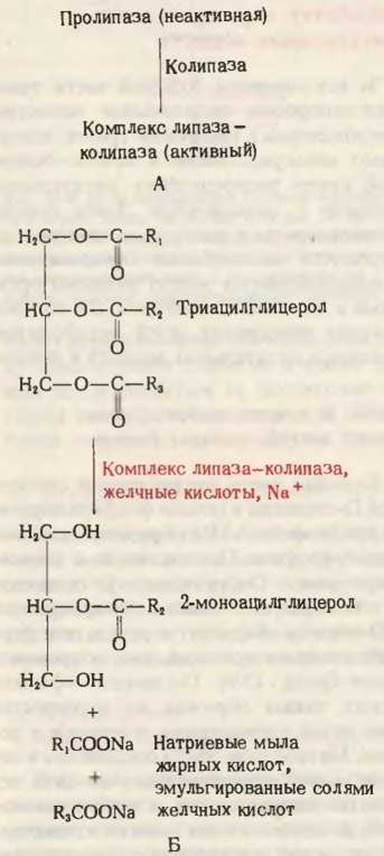
Рис. 24-6. А. Активация липазы. Пролипаза, секретируемая поджелудочной железой, активируется в тонком кишечнике. Колипаза - небольшой белок (мол. масса 10 000), который присоединяется к липазе и стабилизирует фермент. Б. Действие липазы на триацилглицеролы. Липаза катализирует гидролиз триацилглицеролов с высвобождением 2-моноацилглицерола и 1- и 3-ацильных групп в виде омыленных жирных кислот. Протеканию этой реакции способствуют соли желчных кислот, эмульгирующие мыла жирных кислот.
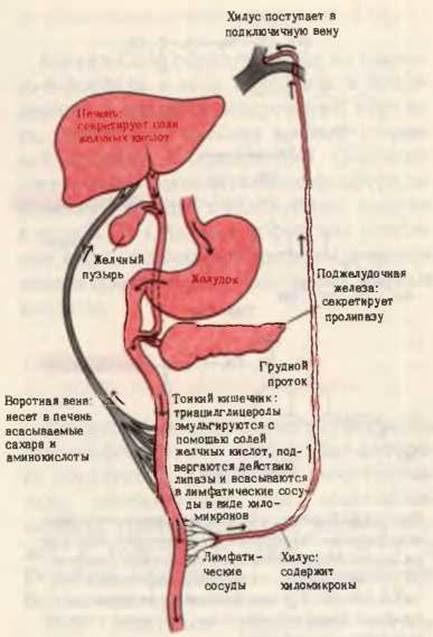
Рис. 24-7. Схематическое изображение основных этапов переваривания и всасывания жиров. Обратите внимание, что желчные кислоты движутся по кругу из печени в тонкий кишечник, затем подвергаются обратному всасыванию, поступают в лимфатические сосуды и почечные вены и возвращаются в печень. Часть желчных кислот обычно теряется в каждом цикле, попадая в кал.
Эмульгированию и перевариванию липидов в тонком кишечнике способствуют соли желчных кислот. Соли желчных кислот человека - это в основном гликохолат натрия и таурохолат натрия, обе они являются производными холевой кислоты (рис. 24-8), которая количественно преобладает среди четырех основных желчных кислот, присутствующих в организме человека. Соли желчных кислот являются мощными эмульгаторами; они поступают из печени в желчь, которая изливается в верхний отдел тонкого кишечника. После завершения всасывания жирных кислот и моноацилглицеролов из эмульгированных капелек жира в нижнем отделе тонкого кишечника происходит обратное всасывание также и солей желчных кислот, способствовавших этому процессу. Они возвращаются в печень и используются повторно. Таким образом, желчные кислоты постоянно циркулируют между печенью и тонким кишечником (рис. 24-7).
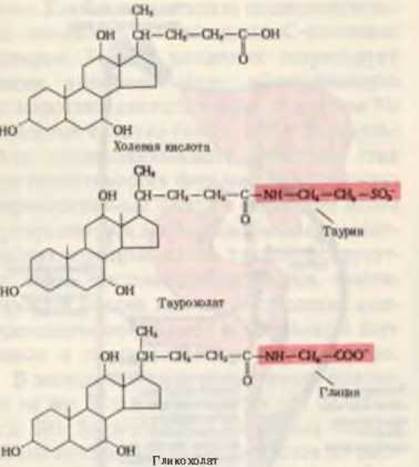
Рис. 24.8. Холевая кислота и ее конъюгированные формы - таурохолат и гликохолат. Благодаря своим амфипатическим свойствам гликохолат и таурохолат - отличные детергенты и эмульгаторы. Группы глицина и таурина (на красном фоне) гидрофильны, тогда как стероидное ядро - гидрофобно.
Желчные кислоты играют исключительно важную роль в усвоении не только триацилглицеролов, но и вообще всех жирорастворимых компонентов пищи. Если желчные кислоты образуются или секретируются в недостаточном количестве, как это имеет место при ряде заболеваний, то непереваренные и непоглощенные жиры появляются в кале. При этом ухудшается всасывание жирорастворимых витаминов A, D, Е и К (разд. 10.13) и может возникнуть пищевая недостаточность витамина А.