Основы биохимии Том 3 - А. Ленинджер 1985
Молекулярные механизмы передачи генетической информации
ДНК: структура хромосом и генов
Уотсон и Крик постулировали модель двойной спирали ДНК
Мы уже видели, как с помощью в высшей степени эффективного метода рентгеноструктурного анализа, основанного на исследовании дифракции рентгеновских лучей, удалось выяснить структуру фибриллярных и глобулярных белков (гл. 7 и 8). При рентгеноструктурном анализе волокон ДНК Розалинд Франклин и Морис Уилкинс получили характерную дифракционную картину (рис. 27-8). На основании этой рентгенограммы был сделан вывод о том, что для полимерных цепей ДНК характерны два типа периодичности вдоль длинной оси: 0,34 и 3,4 нм. Проблема состояла в том, чтобы построить трехмерную модель молекулы ДНК, которая могла бы объяснить не только наличие этих периодичностей, но также открытые Чаргаффом специфические соотношения оснований (А = Т и G = С).
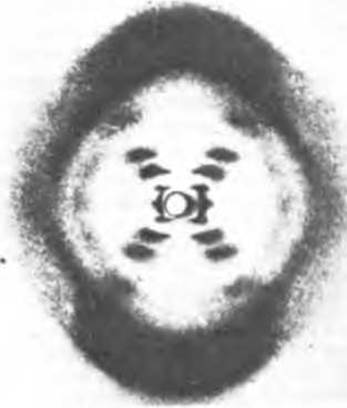
Рис. 27-8. Рентгенограмма ДНК. Крестообразное расположение рефлексов указывает на спиральную структуру молекулы. Сильно затемненные зоны в верхней и нижней частях фотографии соответствуют следующим друг за другом основаниям ДНК.

Рис. 27-9. Уотсон и Крик у одной из своих моделей ДНК (фотография сделана в 1953 г.).
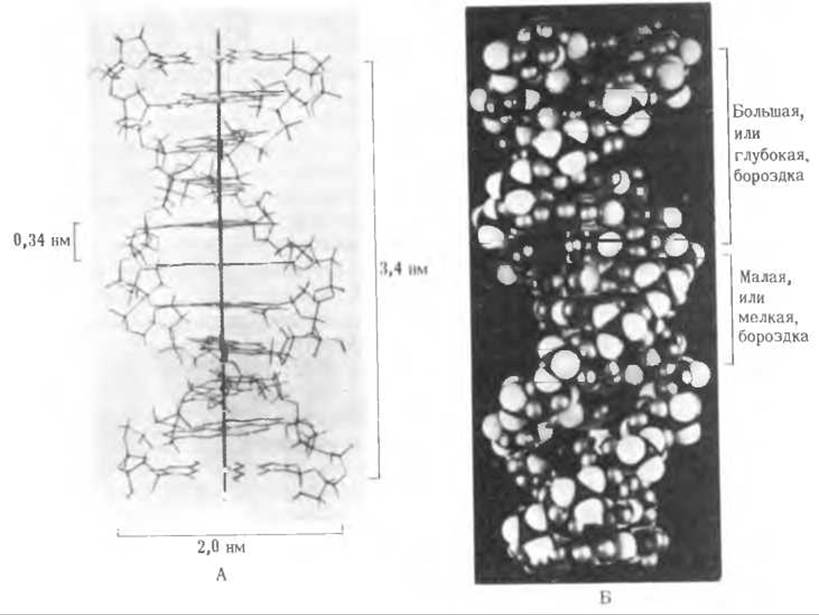
Рис. 27-10. Предложенная Уотсоном и Криком модель структуры ДНК, А. Пространственная проволочная модель. Б. Пространственная модель, в которой отражены объемы атомов.
В 1953 г. американский генетик Джеймс Уотсон и английский физик Фрэнсис Крик как результат своих работ в Кембриджском университете предложили трехмерную модель ДНК, которая объясняла как данные рентгеноструктурного анализа, так и характерную для ДНК парность оснований (рис. 27-10). Модель состоит из двух цепей ДНК, закрученных в спираль вправо вокруг одной и той же оси с образованием двойной спирали. Две цепи в этой спирали антипараллельны, т. е. их 5', 3'-межнуклеотидные фосфодиэфирные мостики направлены в противоположные стороны. Гидрофильные остовы цепей, состоящие из чередующихся остатков дезоксирибозы и отрицательно заряженных фосфатных групп, расположены на внешней стороне двойной спирали и обращены в сторону окружающей ее воды. Гидрофобные пуриновые и пиримидиновые основания обеих цепей уложены стопкой внутри двойной спирали, так что практически плоские молекулы оснований сближены между собой и расположены перпендикулярно длинной оси двойной спирали. Пространственное взаиморасположение цепей приводит к возникновению большой и малой бороздок. Основания одной цепи спарены с находящимися в той же плоскости основаниями другой цепи. Внутри этой структуры точно пригнанными оказываются только определенные пары оснований. Такими соответствующими друг другу парами всегда являются пары пурин-пиримидин, а именно пары А - Т и G - С, т. е. те самые пары, которые образованы основаниями, встречающимися в препаратах ДНК, как это показал Чаргафф, в эквивалентных количествах (табл. 27-3). Более того, основания каждой пары настолько сближены, что между ними возникают водородные связи. Как образуются водородные связи между аденином и тимином и между гуанином и цитозином, показано на рис. 27-11. Важно отметить, что между G и С возникают три водородные связи (G≡C), а между А и Т-только две (А=Т). Другие пары оснований не вписываются в структуру двойной спирали. Пара оснований, составленная из двух пуринов (А и G), слишком велика, чтобы поместиться внутри спирали, имеющей данные параметры, а основания в паре С—Т расположены слишком далеко друг от друга, чтобы образовать стабильные водородные связи. Кроме того, при сохранении своего положения в спирали А не может образовать водородные связи с С, а G - с Т.
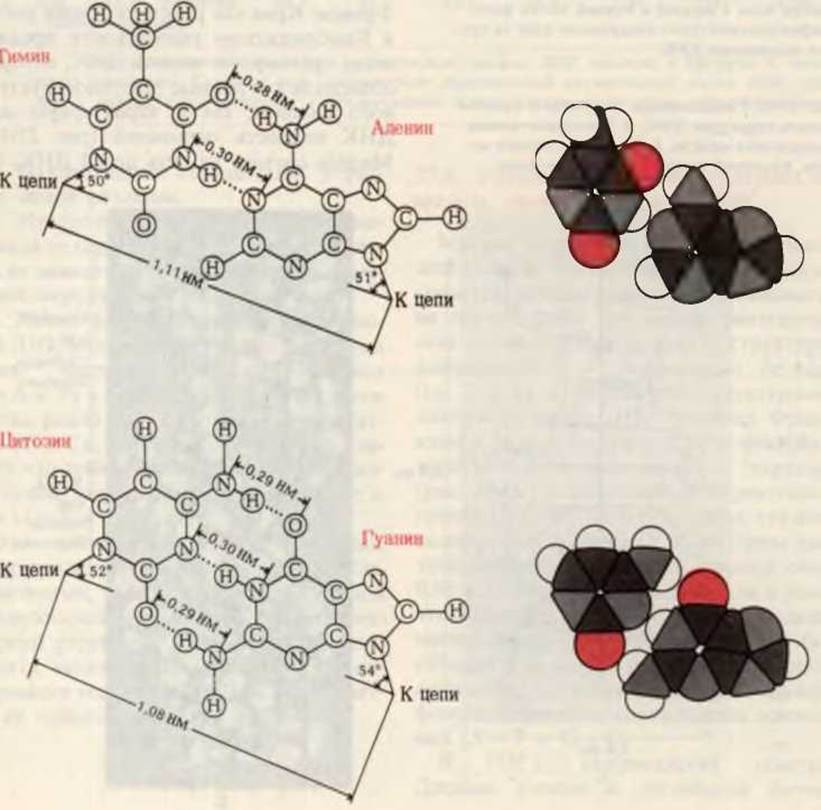
Рис. 27-11. Выполненные в масштабе структурные формулы и пространственные модели соединенных водородными связями оснований в парах аденин-тимин и гуанин-цитозин. Первая из этих пар образована с помощью двух водородных связей, вторая -с помощью трех водородных связей. Пары G-С организованы в пространстве несколько более компактно, чем пары А-Т.
Чтобы объяснить наблюдавшиеся при рентгеноструктурном анализе периодичности, Уотсон и Крик показали с помощью молекулярных моделей, что стопкообразно уложенные внутри двойной спирали основания должны располагаться на расстояниях 0,34 нм друг от друга. Из этих моделей также следовало, что другую периодичность, а именно 3,4 нм можно объяснить тем, что на каждый полный оборот двойной спирали приходится около 10 нуклеотидных остатков (рис. 27-10). Диаметр двойной спирали составляет приблизительно 2 нм. Очень важно обратить внимание на то, что две антипареллельные полинуклеотидные цепи двойной спирали ДНК не идентичны ни по последовательности оснований, ни по нуклеотидному составу, как можно видеть на рис. 27-12. Однако они комплементарны друг другу. Где бы ни появился в одной цепи аденин, напротив него в другой цепи обязательно обнаруживается тимин; точно так же если в одной цепи находится гуанин, то напротив него в другой цепи обязательно присутствует цитозин.
Цепи, образующие двойную спираль ДНК (или дуплекс, как часто называют двойную спираль), удерживаются друг около друга за счет водородных связей между комплементарными основаниями (рис. 27-11) и гидрофобных взаимодействий, благодаря которым уложенные в стопку основания оказываются в значительной степени спрятанными внутрь двойной спирали и защищенными от воды, а сильно полярные остовы полимерных цепей располагаются снаружи и становятся доступными воде. Основной вклад в поддержание стабильности двойной спирали, так же, как в случае третичной структуры глобулярных белков (разд. 8.6), вносят гидрофобные взаимодействия. Отметим, что при pH 7 все фосфатные группы в полярных остовах двойной спирали ионизованы и заряжены отрицательно, так что ДНК представляет собой сильную кислоту.
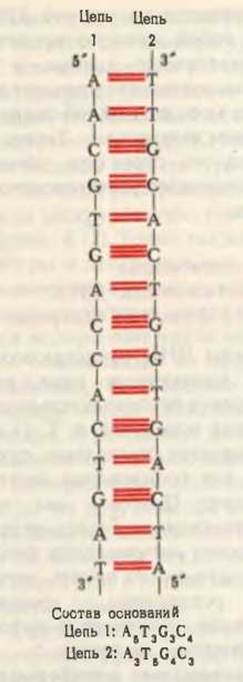
Рис. 27-12. Схематическое изображение комплементарных антипараллельных цепей ДНК согласно модели Уотсона и Крика. Обратите внимание, что цепи отличаются друг от друга по составу оснований, а также по последовательности, если каждую цепь читать в направлении 5'→3'. Отметьте, что соблюдаются равенства, А = Т и G - С.
Многочисленные данные - и химические, и биологические - свидетельствуют о том, что модель двойной спирали ДНК в основном правильна. Теперь посмотрим, как эта структура обеспечивает точное воспроизведение генетической информации.