ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 3. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ - 2017
ЧАСТЬ III. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ
24. ГЕНЫ И ХРОМОСОМЫ
Размер ДНК предлагает нам интересную биологическую загадку, поскольку молекулы ДНК обычно значительно крупнее самих клеток или вирусных частиц (рис. 24-1). Возникает вопрос, как такие крупные молекулы упакованы в клетке. Чтобы ответить на этот вопрос, следует перейти от рассмотрения вторичной структуры ДНК (см. гл. 8) к ее удивительной третичной структуре, лежащей в основе строения хромосом — хранилища генетической информации. В начале главы мы рассмотрим основные элементы хромосом, а затем остановимся на обсуждении их размера и организации. Далее мы обратимся к топологии ДНК и обсудим варианты скручивания и суперскручивания молекул. В заключение мы обсудим взаимодействия ДНК с белками, способствующие компактной укладке хромосом.
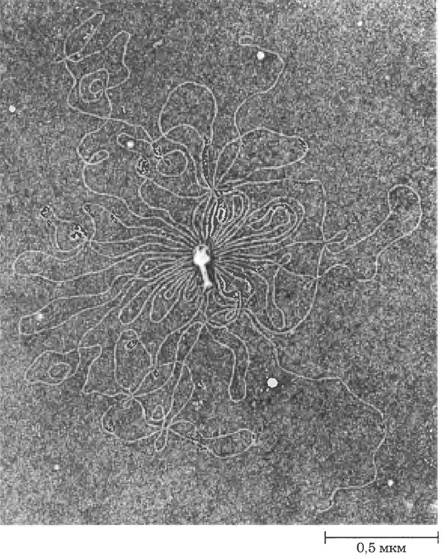
Рис. 24-1. Белковый капсид бактериофага Т2 окружает единственную линейную молекулу ДНК этого фага.
В результате лизиса частиц бактериофага в дистиллированной воде ДНК вышла из капсида и распространилась по поверхности воды. Частица бактериофага Т2 состоит из головки и хвоста, с помощью которого бактериофаг прикрепляется к внешней поверхности бактериальной клетки. Вся ДНК, показанная на этой электронной микрофотографии, обычно содержится внутри головки фага.
24.1. Элементы хромосом
Клеточная ДНК содержит гены и межгенные области; и те, и другие могут выполнять жизненно важные функции. Более сложные геномы, например, геномы эукариот, нуждаются в более сложных уровнях организации хромосом. Мы начнем с рассмотрения различных типов последовательностей ДНК и структурных элементов хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например, цвет глаз. В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент». Позднее эта концепция была расширена до определения «один ген — один полипептид», поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.

Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией. Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации (гл. 28). Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов. Соответствующие механизмы транскрипции и трансляции описаны в гл. 26-28.
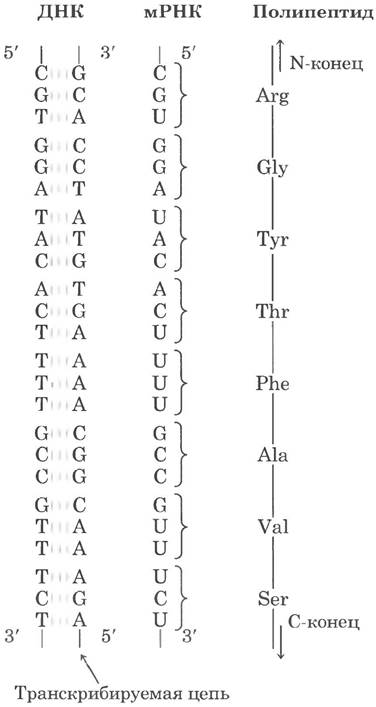
Мы можем приблизительно рассчитать минимальный размер гена, кодирующего средний белок. В гл. 27 подробно рассказано о том, что каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов (рис. 24-2); последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п. н. Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Рис. 24-2 Соответствие между кодирующими участками ДНК, мРНК и аминокислотной последовательностью полипептидной цепи. Триплеты нуклеотидов в ДНК определяют аминокислотную последовательность белка при посредничестве мРНК. Одна из цепей ДНК играет роль матрицы для синтеза мРНК, нуклеотидные триплеты (кодоны) которой комплементарны триплетам ДНК. У некоторых бактерий и многих эукариот кодирующие последовательности прерываются некодирующими участками (так называемыми интронами).
Сколько генов в одной хромосоме? Хромосома прокариота Escherichia coli, чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п. н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Молекулы ДНК гораздо крупнее, чем клеточные или вирусные структуры, в которые они упакованы
Молекулы хромосомной ДНК обычно на много порядков длиннее, чем клетки или вирусные частицы, в которых они содержатся (рис, 24-1; табл. 24-1). Это относится ко всем классам организмов и к вирусам.
Вирусы.
Вирусы не могут жить вне другого организма, вне жизнеспособной клетки. Скорее их можно назвать внутриклеточными паразитами, использующими ресурсы клетки хозяина для размножения. Многие вирусные частицы состоят только из генома (обычно одной молекулы РНК или ДНК), окруженного белковым чехлом.
Геномы почти всех вирусов растений и некоторых вирусов бактерий и животных состоят из РНК. Такие геномы обычно небольшого размера. Например, геномы ретровирусов млекопитающих, таких как ВИЧ. содержат около 9000 нуклеотидов, а бактериофаг Qβ — 4220 нуклеотидов. Геномы обоих вирусов представляют собой одноцепочечную РНК.
Геномы ДНК-содержащих вирусов намного крупнее (табл. 24-1). Многие молекулы ДНК вирусов какую-то часть жизненного цикла находятся в замкнутой кольцевой форме. При репликации вируса в клетке хозяина могут появляться специфические формы вирусных ДНК, называемые репликативными формами; например, многие линейные молекулы ДНК становятся кольцевыми, а одноцепочечные ДНК образуют димеры. Типичным ДНК-содержащим вирусом среднего размера является бактериофаг λ, инфицирующий Е. coli. Репликативная форма ДНК фага λ внутри клеток представлена кольцевой двухцепочечной спиралью. Двухцепочечная ДНК содержит 48 502 п. н., а длина ее контура составляет 17,5 мкм. Геном бактериофага φХ 174 тоже содержит ДНК, но его размер намного меньше; в вирусной частице ДНК представлена одноцепочечной кольцевой молекулой, а двухцепочечная репликативная форма содержит 5386 п. н. Хотя вирусные геномы маленькие, длина их ДНК намного больше, чем размер самих вирусных частиц, содержащих эти молекулы ДНК (табл. 24-1).
Таблица 24-1. Размеры ДНК и вирусных частиц некоторых вирусов бактерий (бактериофагов)
Вирус |
Размер вирусной ДНК, п. н. |
Длина вирусной ДНК, нм |
Длина вирусной частицы, нм |
φХ 174 |
5 386 |
1939 |
25 |
Т7 |
39 936 |
14377 |
78 |
λ |
48 502 |
17460 |
190 |
Т4 |
168 889 |
60800 |
210 |
Примечание. Размер ДНК указав для репликативной (двухцепочечной) формы Длину ДНК оценивали, считая размер пары нуклеотидов равным 3,4 А (см. рис. 8-13 в т. 1).
Бактерии.
В одной клетке Е. coli содержится примерно в 100 раз больше ДНК. чем в частице бактериофага λ. Бактерия Е. coli имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п. н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки Е. coli приблизительно в 650 раз (рис. 24-3). Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами (рис. 24-4; см. также с. 439 в т. 1). Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10 000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах.
Рис. 24-3. Хромосома Е. coli (длиной 1,7 мм) представлена в линейной форме; рядом приведена клетка Е. coli (2 мкм).
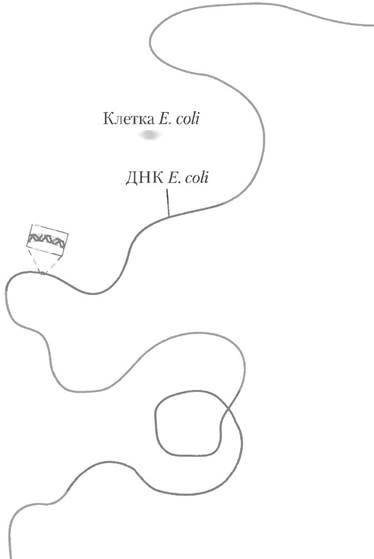
Рис. 24-4. ДНК из лизированной клетки Е. coli. Белыми стрелками отмечены кольцевые молекулы плазмид. Белые и черные пятна — артефакты.
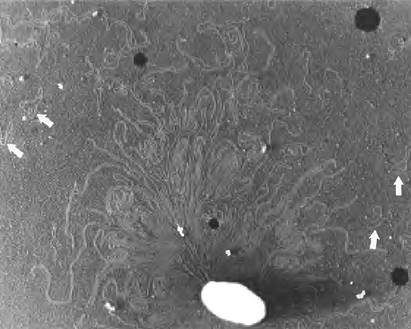
Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача - независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин (см. рис. 6-28 в т. 1). Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
Эукариоты.
В клетке дрожжей, одних из самых маленьких эукариот, в 2,6 раза больше ДНК, чем в клетке Е. coli (табл. 24-2). Клетки плодовой мушки Drosophila, классического объекта генетических исследований, содержат в 35 раз больше ДНК, а клетки человека — примерно в 700 раз больше ДНК, чем клетки Е. coli. Многие растения и амфибии содержат еще больше ДНК. Генетический материал клеток эукариот организован в виде хромосом. Диплоидный набор хромосом (2 n) зависит от вида организма (табл. 24-2). Например, в соматической клетке человека 46 хромосом (рис. 24-5). Каждая хромосома эукариотической клетки, как показано на рис. 24-5, а, содержит одну очень крупную двухспиральную молекулу ДНК. Двадцать четыре хромосомы человека (22 парные хромосомы и две половые хромосомы X и Y) различаются по длине более чем в 25 раз. Каждая хромосома эукариот содержит определенный набор генов.
Таблица 24-2. ДНК, гены и хромосомы некоторых организмов
Общая ДНК, п. н. |
Число хромосомa |
Примерное число генов |
|
Escherichia coli (бактерия) |
4 639 675 |
1 |
4 435 |
Saccharomyces cerevisiae (дрожжи) |
12 080 000 |
16б |
5 860 |
Caenorhabditis elegans (нематода) |
90 269 800 |
12в |
23 000 |
Arabidopsis thaliana (растение) |
119 186 200 |
10 |
33 000 |
Drosophila melanogaster (плодовая мушка) |
120 367 260 |
18 |
20 000 |
Oryza sativa (рис) |
480 000 000 |
24 |
57 000 |
Mus musculus (мышь) |
2 634 266 500 |
40 |
27 000 |
Homo sapiens (человек) |
3 070 128 600 |
46 |
29 000 |
Примечание. Информация постоянно обновляется; для получения более свежей информации обратитесь к сайтам, посвященным отдельным геномным проектам.
а Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом
б Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше наборов таких хромосом.
в Для самок с двумя X хромосомами. У самцов есть X хромосома, но нет Y. т. е. всего 11 хромосом.
Рис. 24-5. Хромосомы эукариот, а — пара связанных и конденсированных сестринских хроматид из хромосомы человека. В такой форме эукариотические хромосомы пребывают после репликации и в метафазе в процессе митоза. б — полный набор хромосом из лейкоцита одного из авторов книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.

Если соединить между собой молекулы ДНК человеческого генома (22 хромосомы и хромосомы X и Y или X и X), получится последовательность длиной около одного метра. Большинство клеток человека диплоидны, поэтому общая длина ДНК таких клеток около 2 м. У взрослого человека примерно 1014 клеток, таким образом, общая длина всех молекул ДНК составляет 2 • 1011 км. Для сравнения, окружность Земли — 4 • 104 км, а расстояние от Земли до Солнца — 1,5 • 108 км. Вот как удивительно компактно упакована ДНК в наших клетках!
В клетках эукариот есть и другие органеллы, содержащие ДНК. это митохондрии и хлоропласты. Молекулы митохондриальной ДНК (мтДНК) намного меньше ядерных хромосом. Размер двухцепочечных кольцевых мтДНК в клетках животных составляет менее 20 000 п. н. (в митохондриях человека молекула мтДНК состоит из 16 569 п. н.). Каждая митохондрия обычно несет от двух до десяти копий молекул мтДНК: их число может возрасти до сотен в некоторых клетках, например, в дифференцирующихся клетках эмбриона. У некоторых организмов (например, трипаносом) каждая митохондрия содержит тысячи копий мтДНК; митохондрии организованы в сложный комплекс, называемый кинетопластом. Размеры мтДНК растительных клеток варьируют от 200 000 до 2 500 000 п. н. Молекулы ДНК хлоропластов (хпДНК) тоже двухцепочечные и кольцевые, их длина колеблется от 120 000 до 160 000 п. н. Выдвигалось множество гипотез относительно происхождения ДНК митохондрий и хлоропластов. Общепризнанная сегодня точка зрения заключается в том, что они представляют собой рудименты хромосом древних бактерий, которые проникли в цитоплазму хозяйских клеток и стали предшественниками этих органелл (см. рис. 1-36 вт. 1). Митохондриальная ДНК кодирует митохондриальные тРНК и рРНК, а также несколько митохондриальных белков. Более 95% митохондриальных белков кодируется ядерной ДНК. Митохондрии и хлоропласта делятся вместе с клетками. ДНК этих органелл реплицируется перед делением клетки и в процессе деления, а затем дочерние молекулы мтДНК попадают в органеллы дочерних клеток.
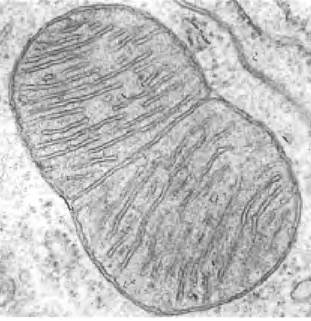
Рис. 24-6. Делящаяся митохондрия. Некоторые митохондриальные белки и молекулы РНК (на фотографии не видны) кодируются одной копией митохондриальной ДНК. Митохондриальная ДНК (мтДНК) реплицируется каждый раз, когда делится митохондрия, что предшествует делению клетки.
Гены и хромосомы эукариот очень сложно организованы
У бактерий многих видов всего одна хромосома, и почти во всех случаях в каждой хромосоме присутствует по одной копии каждого гена. Лишь немногие гены, например, гены рРНК, содержатся в нескольких копиях. Гены и регуляторные последовательности составляют практически весь геном прокариот. Более тою, почти каждый ген строго соответствует аминокислотной последовательности (или последовательности РНК), которую он кодирует (рис. 24-2).
Структурная и функциональная организация генов эукариот гораздо сложнее. Исследование хромосом эукариот, а позднее секвенирование полных последовательностей геномов эукариот принесло много сюрпризов. Многие, если не большинство, генов эукариот обладают интересной особенностью: их нуклеотидные последовательности содержат один или несколько участков ДНК, в которых не кодируется аминокислотная последовательность полипептидного продукта. Такие нетранслируемые вставки нарушают прямое соответствие между нуклеотидной последовательностью гена и аминокислотной последовательностью кодируемого полипептида. Эти нетранслируемые сегменты в составе генов называют интронами, или встроенными последовательностями, а кодирующие сегменты — экзонами. У прокариот лишь немногие гены содержат интроны.
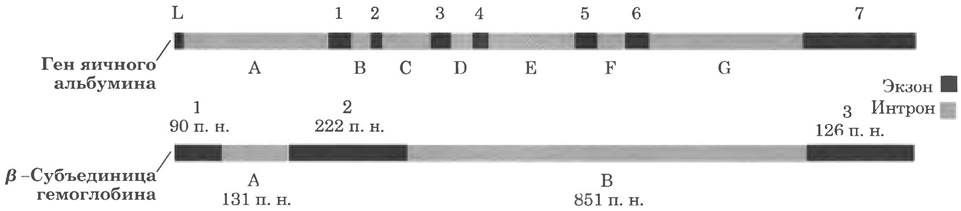
В типичных генах высших эукариот последовательности интронов обычно намного длиннее, чем последовательности экзонов. Например, в гене одной полипептидной цепи яичного белка овальбумина (рис. 24-7) интроны намного длиннее экзонов: семь интронов вместе составляют 85% ДНК гена. В гене β-субъединицы гемоглобина более половины ДНК содержится в единственном интроне. Ген мышечного белка титина чемпион по количеству интронов - их 178. В генах гистонов, по-видимому, интронов нет. В большинстве случаев функция интронов не установлена. В целом, лишь около 1,5% ДНК человека являются «кодирующими», т. е. несут информацию о белках или РНК. Однако с учетом крупных интронов получается, что ДНК человека на 30% состоит из генов.
Рис. 24-7. Интроны в двух эукариотических генах. Ген яичного альбумина содержит семь интронов (от А до G), разделяющих кодирующие последовательности восьми экзонов (L, от 1 до 7). Ген β-субъединицы гемоглобина несет два интрона и три экзона, включая один интрон, содержащий более половины пар нуклеотидов данного гена.
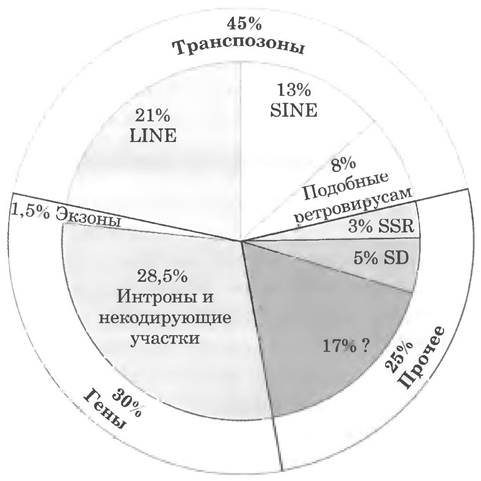
Поскольку гены составляют относительно небольшую долю в геноме человека, значительная часть ДНК остается неучтенной. На рис. 24-8 на круговой диаграмме представлены типы последовательностей в геноме. Большая часть некодирующей ДНК существует в форме повторяющихся последовательностей нескольких типов. Самос удивительное, что примерно половина генома человека состоит из повторяющихся последовательностей подвижных генетических элементов — участков ДНК размером от нескольких сотен до нескольких тысяч пар оснований, которые перемещаются с места на место внутри генома. Эти подвижные генетические элементы (транспозоны) — пример молекулярных паразитов, эффективно расселяющихся внутри генома хозяина. Многие транспозоны несут гены белков, которые катализируют процесс транспозиции, описанный более подробно в гл. 25 и 26. Некоторые транспозоны в геноме человека активны и перемещаются с небольшой частотой, но большинство являются неактивными реликтами, измененными в результате мутаций в процессе эволюции. Хотя чаще всего такие элементы не кодируют функциональные белки или РНК, они сыграли важную роль в эволюции человека, поскольку перемещение транспозонов привело к перераспределению других геномных последовательностей.
Рис. 24-8. Типы последовательностей в геноме человека. На диаграмме представлены три главные составляющие генома — транспозоны (подвижные генетические элементы), гены и смешанные последовательности. Известно четыре основных класса транспозонов (на диаграмме представлены три из них). Длинные рассеянные повторы (LINE) размером от 6 до 8 т. п. н. (1 т. п. н. = 1000 п. н.) обычно содержат несколько генов белков, катализирующих транспозицию. В геноме насчитывается примерно 850 000 LINE. Длина коротких рассеянных повторов (SINE) примерно от 100 до 300 п. н. В геноме человека их содержится около 1,5 млн, из которых более 1 млн составляют элементы Alu, названные так по той причине, что они обычно содержат один сайт рестрикции для эндонуклеазы рестрикции Alul (см. рис. 9-2 в т. 1). В геноме также обнаружено 450 000 копий транпозонов, близких ретровирусам, длиной от 1,5 до 11 т. п. н. Хотя они «захвачены» геномом и не могут перемещаться из клетки в клетку, эволюционно они близки ретровирусам (гл. 26), к которым относится ВИЧ. Еще один класс транспозонов (составляющий <3%, здесь не показан) представлен различающимися по длине фрагментами транспозонов.
Примерно 30% генома составляют последовательности, содержащие гены белков, но только малая часть этой ДНК находится в экзонах (кодирующих последовательностях). К смешанным последовательностям относятся простые короткие повторы (SSR) и повторы крупных сегментов (SD); последние встречаются более чем в одной копии в различных положениях. Как отмечается в гл. 26, почти весь геном в результате транскрипции превращается в последовательность РНК, причем многие участки РНК еще не охарактеризованы. Кроме того, в геноме содержатся остатки транспозонов. которые в процессе эволюции изменились настолько сильно, что их трудно идентифицировать.
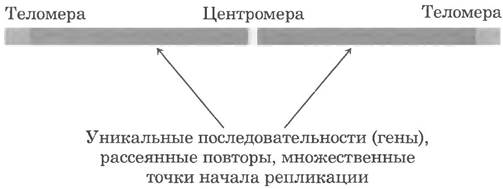
Еще примерно 3% генома человека составляют часто повторяющиеся последовательности, называемые простыми последовательностями ДНК или повторами простых последовательностей (SSR — от англ. simple sequence repeats). Эти короткие последовательности размером обычно менее 10 п. н. иногда повторяются в клетке миллионы раз. Простые последовательности ДНК также называют сателлитной ДНК, поскольку при центрифугировании фрагментов клеточной ДНК в градиенте плотности хлорида цезия из-за необычного состава оснований они часто мигрируют в виде отдельных полос (как спутники, «сателлиты»), сопровождающих остальную ДНК. Простые последовательности ДНК не кодируют белки или РНК. Но в отличие от подвижных генетических элементов сателлитная ДНК может выполнять в клетках человека определенные функции, поскольку многие простые последовательности ДНК локализованы в двух специфических участках эукариотических хромосом - в центромере и теломерах.
Центромера (рис. 24-9) представляет собой последовательность ДНК, к которой в процессе деления клетки прикрепляются белки, связывающие хромосому с митотическим веретеном. Это взаимодействие важно для равномерного и точного разделения наборов хромосом по дочерним клеткам. Были выделены и изучены центромеры Saccharomyces cerevisiae. Важнейшие для функционирования центромер последовательности имеют длину около 130 п. н. и содержат большое количество пар А = Т. Центромеры высших эукариот намного длиннее и, в отличие от дрожжевых, обычно содержат простую последовательность ДНК, состоящую из тысяч тандемных копий одной или нескольких коротких последовательностей из 5-10 п. н. в одинаковой ориентации. Роль простых повторов при функционировании центромер пока точно не установлена.
Рис. 24-9. Важные особенности строения хромосомы дрожжей.
Теломеры (от греч. telos — конец) — последовательности на концах хромосом эукариот, стабилизирующие хромосомы. Теломеры заканчиваются многократно повторяющимися последовательностями вида
(5') (TxGy)n
(3') (АxСy)n
где х и y обычно имеют значения от 1 до 4 (табл. 24-3). Число теломерных повторов n у большинства одноклеточных эукариот составляет от 20 до 100, а у млекопитающих, как правило, превышает 1500. Концы линейной молекулы ДНК не могут полностью реплицироваться обычным способом клеточным репликационным аппаратом (возможно, это одна из причин кольцевой структуры ДНК у бактерий). Теломерные повторы присоединяются к концам эукариотических хромосом в основном с помощью фермента теломеразы (см. рис. 26-39).
Таблица 24-3. Теломерные последовательности
Последовательность теломерных повторов |
|
Homo sapiens (человек) |
(TTAGGG)n |
Tetrahymena thermophila (реснитчатое простейшее) |
(TTGGGG)n |
Saccharomyces cerevisiae (дрожжи) |
((TG)1-3(TG)2-3)n |
Arabidopsis thaliana (растение) |
(TTTAGGG)n |
Для изучения функциональной роли структурных элементов эукариотических хромосом были сконструированы искусственные хромосомы (гл. 9 в т. 1). Для функционирования достаточно стабильной линейной искусственной хромосомы нужны лишь три компонента: центромера, теломеры на каждом конце последовательности и участок, в котором происходит инициация репликации. Искусственные хромосомы дрожжей (YAC; см. рис. 9-7) были созданы как инструмент для биотехнологических исследований. Для лечения генетических заболеваний с помощью соматической генной терапии были разработаны искусственные хромосомы человека (НАС; см. доп. 9-2).
Краткое содержание раздела 24.1 Элементы хромосом
■ Ген - участок хромосомы, который несет информацию о функциональном полипептиде или молекуле РНК. Наряду с генами хромосомы содержат разнообразные регуляторные последовательности, участвующие в репликации, транскрипции и других процессах.
■ Геномная ДНК и геномная РНК вирусов обычно на несколько порядков длиннее содержащих их клеток или вирусных частиц.
■ Многие гены эукариот (но лишь некоторые гены бактерий и архей) прерываются некодирующими последовательностями — нитронами. Кодирующие участки, разделенные нитронами, называют экзонами.
■ Менее трети геномной ДНК человека составляют гены. В остальной ДНК много повторяющихся последовательностей различных типов. Паразитирующие нуклеиновые кислоты, известные как транспозоны, составляют примерно половину генома человека.
■ Хромосомы эукариот содержат две важные повторяющиеся последовательности ДНК со специфической функцией: центромеры (участки прикрепления митотического веретена) и теломеры, расположенные на концах хромосом.