ОСНОВЫ БИОХИМИИ ЛЕНИНДЖЕРА - ТОМ 3. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ - 2017
ЧАСТЬ III. ПУТИ ПЕРЕДАЧИ ИНФОРМАЦИИ
24. ГЕНЫ И ХРОМОСОМЫ
24.3. Структура хромосом
Хромосомами называют молекулы нуклеиновых кислот, которые служат хранилищем генетической информации у вирусов, бактерий, клеток прокариот или органелл эукариот. Этот термин также относится к интенсивно окрашиваемым тельцам, которые видны в световой микроскоп, после обработки красителями в ядрах эукариотических клеток.
Хроматин состоит из ДНК и белков
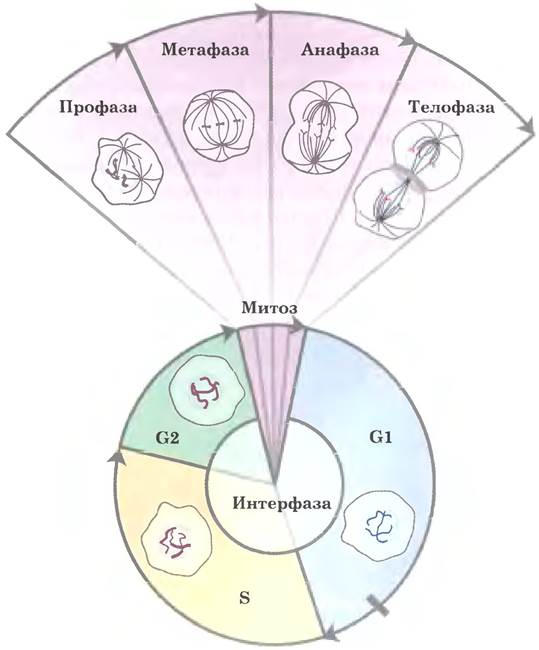
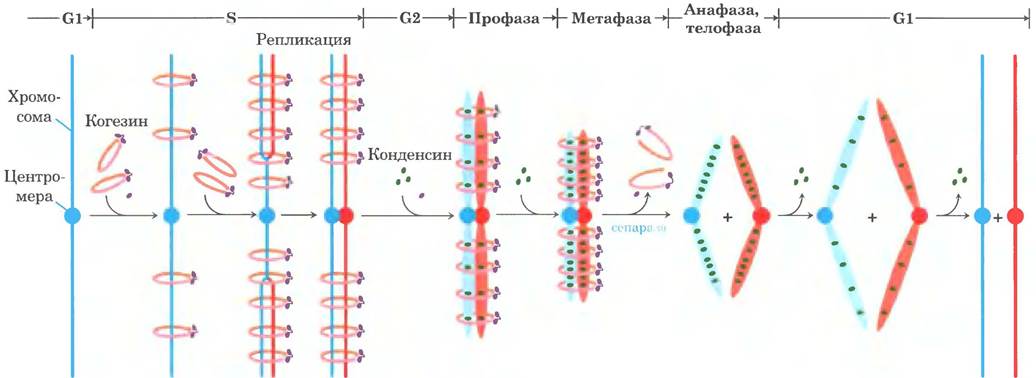
В клеточном цикле эукариот (см. рис. 12-43 в т. 1) структура хромосом изменяется удивительным образом (рис. 24-25). В неделящихся клетках эукариот (стадия G0) и на стадии интерфазы при делении (G1, S и G2) материал хромосом, хроматин, аморфен и, вероятно, хаотически распределен по центру ядра. В фазе S интерфазы ДНК в таком аморфном виде реплицируется, при этом каждая хромосома образует две сестринские хромосомы (они называются сестринскими хроматидами), которые остаются связанными друг с другом после завершения репликации. Хромосомы становятся намного более конденсированными во время профазы митоза и принимают характерную форму, которая зависит от вида организма (рис. 24-5).
Рис. 24-25. Изменения структуры хромосомы в ходе клеточного цикла эукариот. Относительная длительность фаз изображена здесь только для удобства. Длительность каждой фазы различна в разных типах клеток и при разных условиях роста (для одноклеточных организмов) или состояния метаболизма (для многоклеточных). Самой короткой обычно бывает фаза митоза. Как схематично показано на диаграмме, в интерфазе клеточная ДНК деконденсирована. В интерфазе (см. рис. 12-43 в т. 1) можно отдельно выделить фазу G1 (от англ. дар — промежуток), фазу S (синтез), когда происходит репликация ДНК, и фазу G2, во время которой реплицированные хромосомы (хроматиды) сходятся друг с другом. В митозе можно выделить четыре фазы. Конденсация ДНК происходит в профазе. Во время метафазы конденсированные хромосомы выстраиваются парами в центре веретена деления. Каждая хромосома в каждой паре хромосом связана с веретеном деления, направленным к полюсу клетки, через микротрубочки, которые расположены между веретеном и центромерой. Сестринские хроматиды разделяются в анафазе, каждая из них движется к полюсу, с которым она соединена веретеном. После завершения деления клетки хромосомы деконденсируются, и цикл начинается заново.
Хроматин состоит из нитей, содержащих белок и ДНК примерно в равной пропорции (по массе) и небольшое количество РНК. Молекула ДНК в хроматине очень плотно связана с белками гистонами, которые упаковывают и упорядочивают ДНК в структурные единицы — нуклеосомы (рис. 24-26). В хроматине также обнаружено много негистоновых белков — одни из них помогают поддерживать структуру хромосом, а другие регулируют экспрессию специфических генов (гл. 28). Начиная с нуклеосом, ДНК эукариот образует более высокоорганизованные структуры, которые в итоге формируют компактную хромосому, видимую в световом микроскопе. Рассмотрим структуру хромосом эукариот и сопоставим ее с упаковкой ДНК в клетках бактерий.
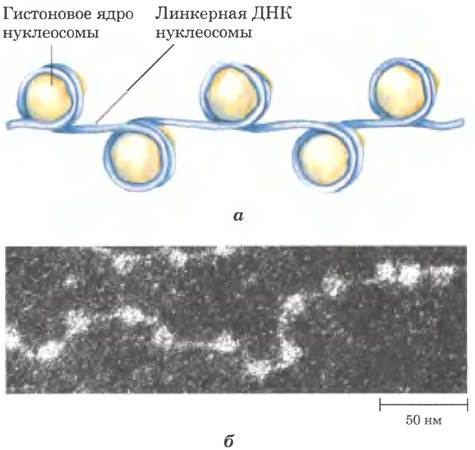
Рис. 24-26. Нуклеосомы. Разделенные равными промежутками нуклеосомы состоят из гистоновых комплексов, связанных с ДНК. а — схематичное изображение; 6 — электронная микрофотография.
Гистоны — небольшие основные белки
Гистоны, с молекулярной массой от 11 000 до 21 000, обнаружены во всех клетках эукариот; эти белки отличаются высоким содержанием основных аминокислот аргинина и лизина (которые вместе составляют четверть всех аминокислот). Все эукариотические клетки содержат пять основных классов гистонов, различающихся молекулярной массой и аминокислотным составом (табл. 24-4). Гистоны Н3 имеют почти идентичную аминокислотную последовательность в клетках всех эукариот, как и гистоны Н4, что указывает на консервативность их функций. Например, молекулы гистонов H4 гороха и коровы различаются только двумя аминокислотными остатками из 102; молекулы гистонов H4 человека и дрожжей различаются восемью остатками. Последовательности гистонов H1, Н2А и Н2В у разных видов эукариот менее схожее.
Таблица 24-4. Классы гистонов и их свойства
Молекулярная масса |
Число аминокислотных остатков |
Доля основных аминокислот (% от общего числа) |
||
Гистон |
Lys |
Arg |
||
Н1* |
21 130 |
223 |
29,5 |
11,3 |
Н2А* |
13 960 |
129 |
10,9 |
19,3 |
Н2В* |
13 774 |
125 |
16,0 |
16,4 |
Н3 |
15 273 |
135 |
19,6 |
13,3 |
Н4 |
11236 |
102 |
10,8 |
13,7 |
* Размеры этих гистонов слегка различаются у разных видов. Приведенные данные относятся к гистонам быка.
Гистоны каждого класса подвергаются ферментативной модификации — метилированию, ADР-рибозилированию, фосфорилированию, гликозилироваиию или ацетилированию. Такие модификации влияют на суммарный электрический заряд, форму и другие свойства гистонов, на структурные и функциональные свойства хроматина, а также важны для регуляции транскрипции (гл. 28).
Кроме того, некоторые гистоны эукариот существуют в виде нескольких вариантов, особенно это касается гистонов Н2А и Н3, которые более подробно описаны ниже. Эти гистоны и их модификации играют в метаболизме ДНК особую роль.
Нуклеосомы — основные структурные единицы хроматина
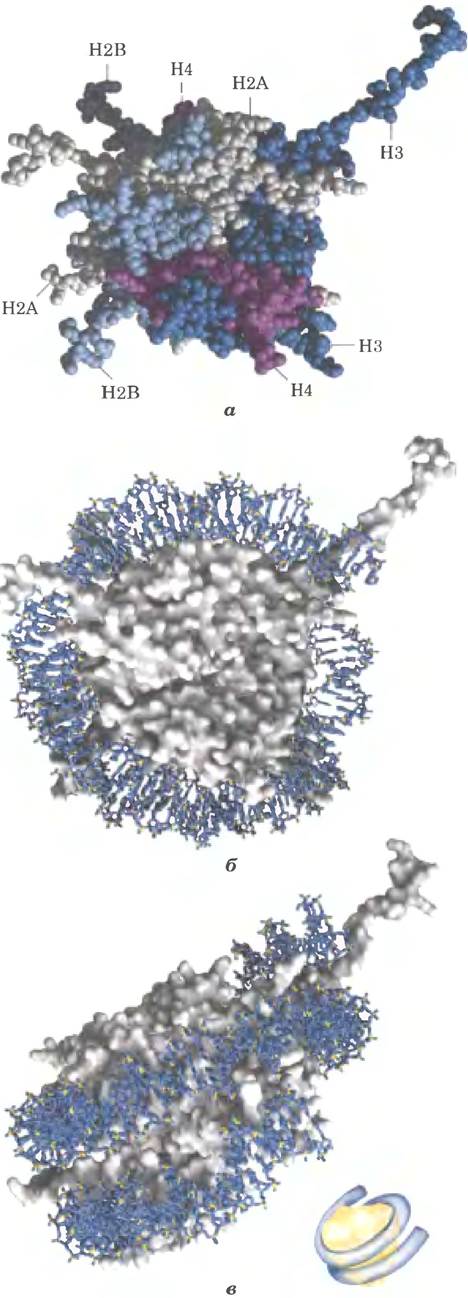
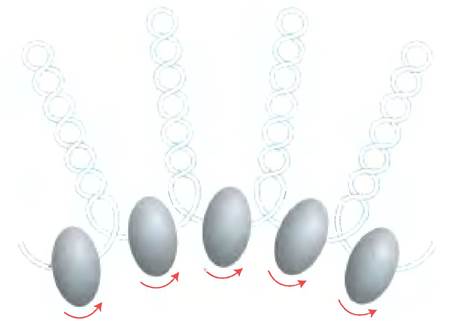
Эукариотические хромосомы, изображенные на рис. 24-5, представляют собой компактную укладку молекулы ДНК длиной примерно 105 мкм в ядре клетки, диаметр которого обычно составляет от 5 до 10 мкм. Такая компактность достигается благодаря многоуровневой организации укладки. При частичном нарушении укладки хромосомы обнаруживается структура, в которой ДНК прочно связана с глобулами (шариками) белка, часто разделенными регулярными промежутками (рис. 24-27). Шарики в этих «бусах» — это гистоны в комплексе с ДНК. Шарик и нить ДНК, соединяющая его со следующим шариком, образуют нуклеосому — основную структурную единицу, на которой происходит дальнейшая укладка хроматина. Глобула каждой нуклеосомы содержит восемь гистонов: по две копии Н2А, Н2В, Н3 и Н4. Фрагменты ДНК между шариками представляют собой повторяющиеся единицы, обычно длиной 200 п. н., из которых 146 п. н. плотно обмотаны вокруг гистонового ядра из восьми частей, а оставшаяся часть ДНК соединяет глобулы между собой. Гистон Н1 взаимодействует с соединяющей (линкерной) ДНК. Неглубокая обработка хроматина расщепляющими ДНК ферментами вызывает в первую очередь деградацию линкерной ДНК; в результате происходит высвобождение гистоновых частиц, содержащих 146 п. н. связанной ДНК, которая была защищена от расщепления. Исследователи кристаллизовали полученные таким образом нуклеосомные частицы и с помощью рентгеноструктурного анализа установили, что ДНК обмотана вокруг восьми молекул гистонов в виде левозакрученной соленоидной сверхспирали (рис. 24-27).
Рис. 24-27. ДНК обвивается вокруг ядра нуклеосомы. а — шаровая модель белков нуклеосомной частицы из африканской лягушки Xenopus laevis; разными цветами показаны разные гистоны (PDB ID 1A0I). б и в — вид сверху и вид сбоку на кристаллическую структуру нуклеосомы (контур поверхности показан серым цветом) с обвитой связанной ДНК (синяя) длиной 146 п. н. Молекула ДНК связывается в виде левозакрученной соленоидной сверхспирали, опоясывающей гистоновый комплекс 1,8 раз. Для наглядности представлено схематичное изображение нуклеосомы (в — внизу справа).
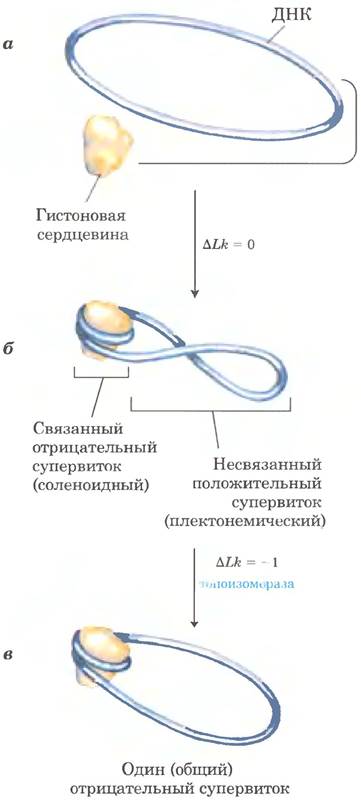
Детальное изучение данной структуры объяснило, почему ДНК эукариот частично раскручивается даже в отсутствие ферментов в эукариотических клетках, осуществляющих частичное раскручивание ДНК. Вспомним, что соленоидное скручивание ДНК в нуклеосомах - одна из двух форм сверхспирали, которую может принимать частично раскрученная (отрицательно сверхспирализованная) ДНК. Плотное обматывание ДНК вокруг гистонового ядра требует удаления из ДНК примерно одного витка. Когда белковая сердцевина нуклеосомы связывается in vitro с релаксированной замкнутой кольцевой ДНК. происходит отрицательная сверхспирализация. Поскольку такой процесс связывания не разрывает ДНК и не изменяет порядок зацепления, формирование отрицательной соленоидной сверхспирали должно компенсироваться образованием положительной сверхспирали на участке несвязанной ДНК (рис. 24-28). Как упоминалось ранее, эукариотические топоизомеразы могут вызывать релаксацию положительной сверхспирали. Релаксация несвязанной положительно сверхспирализованной ДНК приводит к фиксированию отрицательно сверхснирализованной ДНК (при ее связывании с гистоновой сердцевиной) и выражается в общем уменьшении порядка зацепления. Было показано, что топоизомеразы необходимы для сборки хроматина из очищенных гистонов и замкнутой кольцевой ДНК in vitro.
Рис. 24-28. Сборка хроматина, а — релаксированная замкнутая кольцевая ДНК. б — при связывании гистоновой сердцевины и формировании нуклеосомы возникает один отрицательный сверхвиток. Если ни в одной из цепей нет разрывов, где-нибудь в ДНК возникает положительный сверхвиток (∆Lk = 0). в — релаксация такого положительного сверхвитка топоизомеразой оставляет один отрицательный сверхвиток (∆Lk = -1).
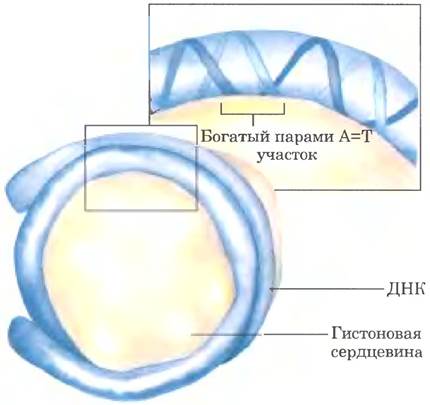
Еще одним фактором, влияющим на связывание ДНК с гистонами в нуклеосомах, является последовательность ДНК. Гистоновая сердцевина связывается не с любым, а с определенным участком ДНК. Принцип выбора такого участка до конца не ясен, но в некоторых случаях, вероятно, он определяется количеством пар А = Т в том участке спирали ДНК, где она соприкасается с гистонами (рис. 24-29). Сужение малой бороздки спирали в этих точках способствует плотной упаковке ДНК вокруг гистонового ядра; сочетание двух или трех пар нуклеотидов А = Т увеличивает вероятность такого сужения. Особенно охотно нуклеосомы связываются с такими последовательностями, в которых динуклеотиды АА, АТ или ТТ встречаются с интервалом 10 п. н., и именно таким свойством обладают до 50% последовательностей, связывающихся с гистонами in vivo.
Рис. 24-29. Расположение нуклеосомы, при котором оптимально используются пары нуклеотидов А = Т в участках соприкосновения гистоновой сердцевины с малой бороздкой спирали ДНК.
Для правильного расположения некоторых нуклеосомных частиц на ДНК необходимы другие белки. У некоторых организмов определенные белки связываются со специфической последовательностью ДНК, способствуя формированию поблизости нуклеосомной частицы. В ходе репликации или в других процессах, требующих временного перемещения нуклеосом, нуклеосомы остаются связанными с ДНК. Это связывание осуществляется ступенчатым образом. Сначала происходит связывание тетрамера, образованного двумя гистонами Н3 и двумя гистонами Н4, вслед за которыми связываются димеры Н2А Н2В. Связывание нуклеосом с хромосомами после репликации хромосом происходит при участии белкового комплекса RCAF (от англ. replication-coupling assembly factor). Комплекс RCAF состоит из ацетилированиых гистонов Н3 и Н4, грехсубъединичного белка, называемого фактором сборки хроматина-1 (англ. chromatin assembly factor 1, CAF1), и белка ASF1 (от англ. antisilencingfactor 1). Механизм связывания нуклеосом изучен не полностью, хотя известно, что элементы комплекса RCAF непосредственно взаимодействуют с элементами репликационного аппарата. Когда после репарации ДНК или другого процесса возникает необходимость сборки нуклеосом, RCAF заменяется другим специализированным белковым комплексом, опосредующим связывание. В некоторых случаях факторы обмена гистонов позволяют осуществлять замену основных гистонов на другие варианты. Правильное расположение этих вариантов гистонов очень важно. Например, при отсутствии одного из вариантов гистонов эмбрионы мыши погибают на ранних стадиях развития (дон. 24-2). Точное расположение нуклеосомных частиц также может иметь значение для экспрессии некоторых эукариотических генов (гл. 28).
Нуклеосомы образуют структуры с более сложной организацией
Закручивание вокруг нуклеосомной частицы уменьшает длину ДНК примерно в семь раз. Но в целом в хромосоме длина ДНК сокращается более чем в 10 000 раз, следовательно, существуют и другие уровни организации ДНК. Очень мягкие методы выделения хромосом позволяют обнаружить, что ядра нуклеосом организованы в структуры, называемые 30-нм фибриллами (рис. 24-30). Для такой упаковки требуется одна молекула гистона Н1 на нуклеосому. Однако участки такой укладки прерываются участками ДНК, связанными со специфическими (негистоновыми) белками. Кроме того, наличие 30-нм фибрилл зависит от интенсивности транскрипции участка ДНК. Участки активной транскрипции генов находятся в менее организованном состоянии и содержат очень мало гистона Н1 (или вовсе его не содержат).
Рис. 24-30.30-нм фибриллы — более высокий уровень организации нуклеосом. а — схема предполагаемого строения фибриллы, б — электронная микрофотография.
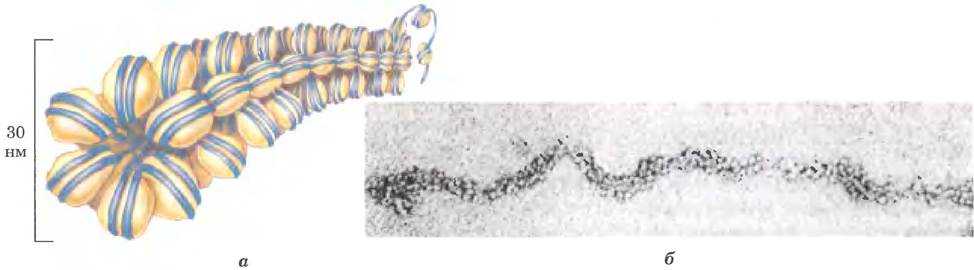
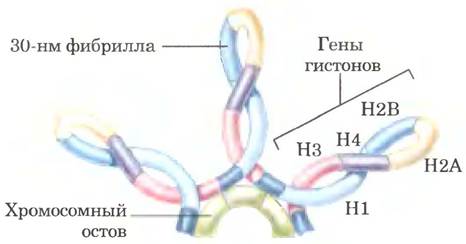
30-нм фибриллы представляют собой второй уровень организации хроматина, обеспечивающий 100-кратное сокращение длины ДНК. Более высокие уровни организации ДНК пока не исследованы; вероятно, определенные участки ДНК связаны с хромосомным остовом (рис. 24-31). Ассоциированные с остовом участки разделены петлями ДНК размером от 20 до 100 т. и. н. ДНК в этой петле может содержать набор родственных генов. Например, у дрозофил полный набор генов, кодирующих гистоны, сосредоточен в виде кластеров в петлях, которые присоединены к хромосомному остову (рис. 24-32). Сам осьов также содержит несколько белков, особенно в большом количестве гистон Н1 (расположенный на внутренней стороне волокна) и топоизомеразу II. Присутствие топоизомеразы II еще раз указывает на связь между частичным раскручиванием ДНК и структурой хроматина. Топоизомераза II настолько важна для поддержания структуры хроматина, что ингибиторы этого фермента могут быстро уничтожить делящиеся клетки. Несколько химиотерапевтических препаратов являются ингибиторами топоизомеразы II - фермент может инициировать разрыв нитей, но не позволяют ему зашить разрыв.
Рис. 24-31. Частично расплетенная хромосома человека; видны многочисленные петли ДНК, прикрепленные к хромосомному остову.
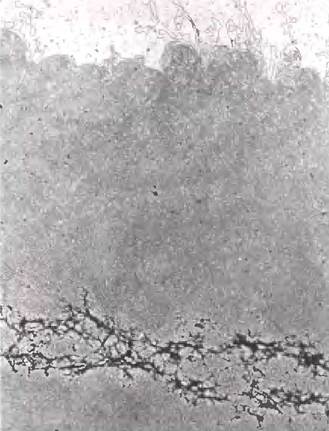
Рис. 24-32. Петли хромосомной ДНК, прикрепленные к хромосомному остову. Молекулы ДНК в петлях упакованы в виде 30-нм фибрилл, поэтому петли — следующий уровень организации хромосом. Петли часто содержат группы генов с родственными функциями. Полные наборы генов гистонов, как показано на данной схеме, образуют кластеры в петлях такого рода. В отличие от большинства других гены гистонов во многих геномах эукариот присутствуют в большом количестве копий.
Дополнение 24-2. МЕДИЦИНА. Эпигенетика, структура нуклеосом и варианты гистонов
Информация, которая передается от одного поколения к другому (дочерним клеткам при делении клеток или детям от родителей), но не кодируется в ДНК, называется эпигенетической. Большая часть этой информации содержится в виде ковалентных модификаций гистонов и/или в виде размещения вариантов гистонов в хромосомах.
Участки хроматина, в которых происходит активная экспрессия генов (транскрипция), обычно находятся в неконденсированной форме и называются эухроматином. В этих участках гистоны Н3 и Н2А часто заменены на варианты гистонов Н3.3 и H2AZ соответственно (рис. 1). Комплексы, осуществляющие связывание ДНК с нуклеосомами, содержащими варианты гистонов, сходны с теми, что отвечают за связывание с нуклеосомами. содержащими основные гистоны. Связывание нуклеосом. содержащих гистон Н3.3, осуществляется комплексом, в котором фактор сборки хрома- тина-1 (CAF1) заменен белком HIRA (название происходит от класса белков HIR, от англ. histone repressor). Белки CAF1 и HIRA можно рассматривать в качестве гистоновых шаперонов. помогающих обеспечить правильную сборку и размещение нуклеосом. Гистон Н3.3 отличается от Н3 лишь четырьмя аминокислотными остатками, однако эти остатки играют ключевую роль в связывании гистона.
Рис. 1. Известно несколько вариантов гистонов Н3, Н2А и Н2В. Здесь показаны основные гистоны и несколько известных вариантов. Выделены участки метилирования остатков Lys/Arg и фосфорилирования остатков Ser. Участок HFD (от англ. histone-fold domain) — структурный домен, присутствующий во всех основных гистонах.
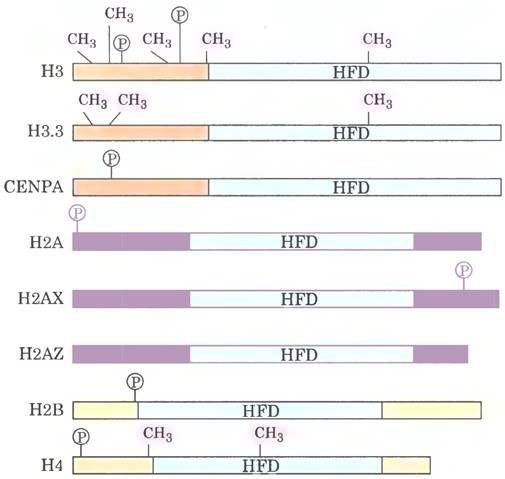
Подобно гистону Н3.3, гистон H2AZ связан с определенным связывающим комплексом и обычно ассоциирован с активно транскрибирующимися участками хроматина. Включение H2AZ стабилизирует нуклеосомный октамер, но препятствует некоторым кооперативным взаимодействиям между нуклеосомами, необходимым для формирования компактной структуры хромосомы. Это приводит к образованию хромосомы с более открытой структурой, что облегчает экспрессию генов в области H2AZ. Ген, кодирующий H2AZ, играет важную роль в развитии млекопитающих. У дрозофил отсутствие H2AZ препятствует развитию организма после стадии личинки.
Другой вариант гистона Н2А — Н2АХ связан с репарацией ДНК и генетической рекомбинацией. У мышей отсутствие Н2АХ приводит к геномной нестабильности и бесплодию у самцов. Небольшие количества ШАХ, по-видимому, распределены по всему геному. При двухцепочечном разрыве ДНК находящиеся поблизости молекулы Н2АХ фосфорилируются по остатку Ser139 на С-конце белка. Если блокировать фосфорилирование экспериментальным путем, ингибируется образование белковых комплексов, необходимых для репарации ДНК.
Вариант гистона Н3, известный как CENPA, связан с повторяющимися последовательностями ДНК в центромерах. Хроматин в области центромеры содержит гистоновые шапероны CAF1 и HIRA; по-видимому, оба белка участвуют в связывании нуклеосом, содержащих CENPA. Удаление гена CENPA у мышей приводит к летальному исходу.
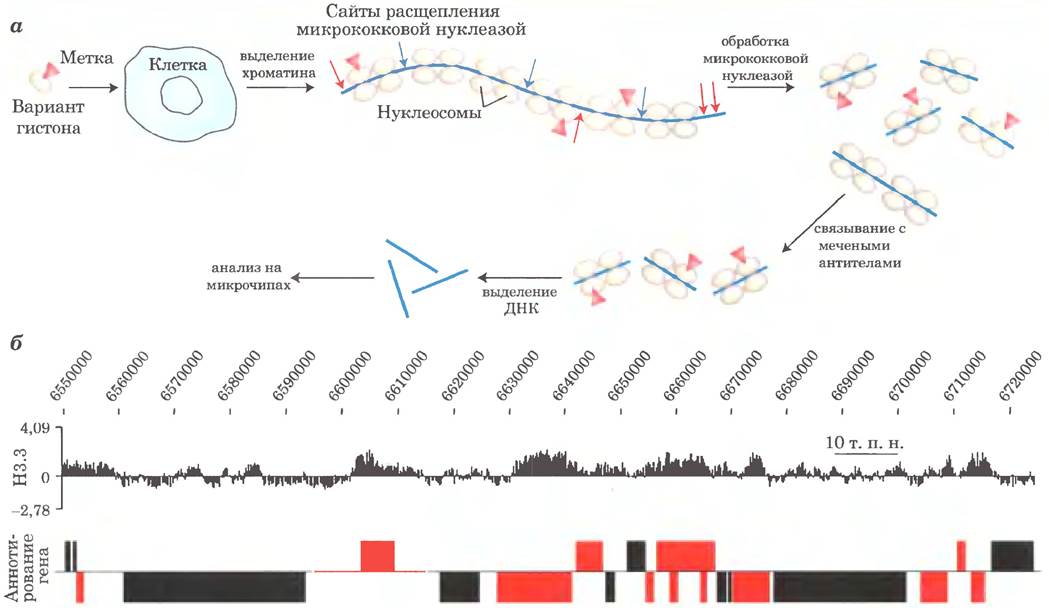
Для изучения функций и расположения вариантов гистонов можно применять методы геномики. Одним из полезных методов является иммунопреципитация хроматина (СhIP, от англ. chromatin immunoprecipitatiori). Суть метода состоит в осаждении нуклеосом, содержащих определенный вариант гистона, при связывании этого варианта со специфическими антителами. В принципе нуклеосомы можно изучать отдельно от ДНК, но чаще при исследовании связанную ДНК сохраняют, что позволяет определить участок связывания нуклеосомы. Молекулу ДНК можно пометить и использовать в качестве зонда в методе анализа на микрочипах (см. рис. 9-22 в т. 1) для получения карты геномной последовательности, с которой связывается каждая конкретная нуклеосома. Этот метод часто называют иммунопреципитацией на чипе (СhIР on chip; рис. 2).
Рис. 2. Метод иммунопреципитации хроматина на микрочипе позволяет определить последовательности генома, с которыми связывается тот или иной вариант гистона, а — вариант гистона с меткой-эпитопом (белком или химической структурой, распознаваемой антителом; см. гл. 5 и 9 в т. 1) вводят в определенный тип клеток, где он встраивается в нуклеосомы. (В некоторых случаях в таких метках нет нужды, поскольку существуют антитела, связывающиеся непосредственно с модифицированным участком изучаемого гистона.) Затем из клеток выделяют хроматин и расщепляют его микрококковой нуклеазой. Связанная с нуклеосомами ДНК защищена от расщепления, однако линкерная ДНК расщепляется, что приводит к высвобождению сегментов ДНК, связанных с одной или двумя нуклеосомами. При добавлении антител происходит селективное осаждение нуклеосом, содержащих меченый вариант гистона. Из осадка экстрагируют связанную с этими нуклеосомами ДНК, помечают ее и используют в качестве зонда для анализа на микрочипах, содержащих геномные последовательности данного типа клеток. 6 — гибридизация на микрочипе позволяет обнаружить последовательности ДНК, связанные с нуклеосомами с определенным вариантом гистона. В данном примере (верхняя панель) представлена гибридизация ДНК, связанной с гистоном Н3.3 в небольшом участке генома дрозофилы. Аннотированные гены данного участка генома представлены на нижней панели (широкие полосы). С гистоном Н3.3 связываются участки ДНК, расположенные вблизи активно транскрибирующихся генов (красные полосы).
Варианты гистонов, а также многие ковалентные модификации гистонов, позволяют определить функции хроматина. Они размечают хроматин, облегчая или подавляя специфические функции, такие как расхождение хромосом, транскрипция и репарация ДНК. Модификации гистонов не исчезают при делении клеток или при мейозе, и таким образом они становятся частью информации, передаваемой от поколения к поколению во всех эукариотических организмах.
Получены доказательства еще более сложной организации хромосом эукариот, причем каждый уровень организации значительно повышает компактность упаковки. Одна из моделей, объясняющих компактность структуры ДНК, показана на рис. 24-33. Более высокоорганизованные структуры хроматина, вероятно, различаются в разных хромосомах, в разных участках одной хромосомы и в разные моменты жизненного цикла клетки. Ни одна модель не может адекватно описать все эти структуры.
Рис. 24-33. Компактная упаковка ДНК в хромосоме эукариот. Модель демонстрирует уровни организации ДНК эукариот, которые могут объяснить компактную упаковку хромосом. В клетке уровни более высокой организации (выше уровня 30-нм фибрилл), скорее всего, не столь единообразны, как показано на данной схеме.
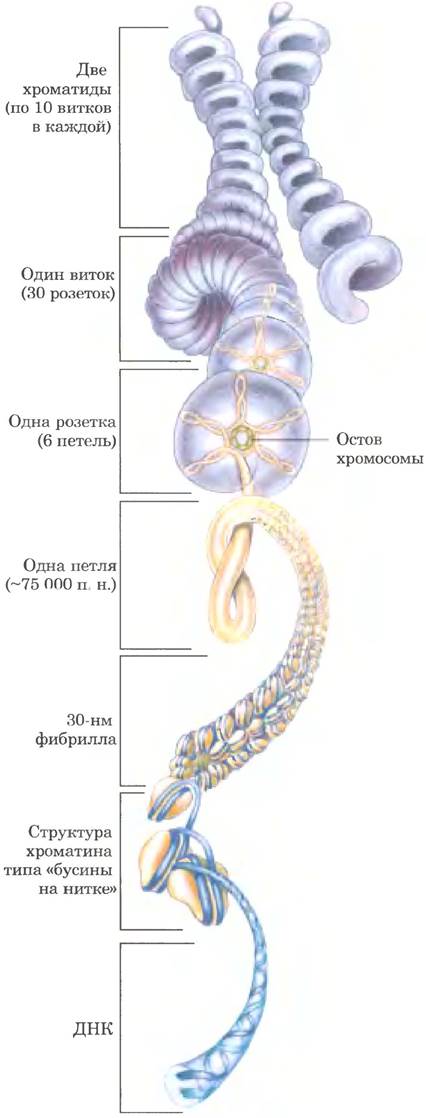
Тем не менее принцип ясен: компактность ДНК эукариотических хромосом обеспечивается укладкой витка на виток, витка на виток...
Структура конденсированных хромосом поддерживается SMC-белками
Наряду с гистонами и топоизомеразами существует третий важный класс белков хроматина — это так называемые SMC-белки (от англ. structural maintenance of chromosomes — белки, поддерживающие структуру хромосом). Первичная структура SMC-белков состоит из пяти различимых доменов (рис. 24-34, а). Концевые глобулярные домены N и С (на N- и С-конце), каждый из которых содержит часть активного центра, в котором происходит гидролиз АТР, соединены с двумя α-спиральными участками (см. рис. 4-10 в т. 1), между которыми располагается шарнирный домен. Белки этого класса обычно образуют димерный V-образный комплекс, соединяясь друг с другом через шарнирный домен (рис. 24-34, б, в). Один N- и один С-домен сближаются, образуя функциональный центр гидролиза АТР на каждом свободном конце V-образного комплекса.
Рис. 24-34. Структура SMC-белков, а — пять доменов первичной структуры SMC. Буквами N и С обозначены N- и С-концевые последовательности соответственно, б — каждый полипептид сложен таким образом, что два спиральных домена закручены относительно друг друга, так что N- и С-домены соединяются, образуя полноценный ATP-связывающий сайт. Два таких домена связываются шарнирной областью с образованием V-образного димера. в — электронная микрофотография SMC-белков из Bacillus subtilis. г — когезины состоят из пар белков SMC1 и SMC3, а конденсины — из пар SMC2-SMC4. Эти эукариотические SMC-белки связываются с клейзином и некоторыми другими регуляторными белками (не показаны), д - гидролиз АТР может служить для открытия и закрытия АТРазного домена SMC-димера, который остается связанным с клейзином и другими белками (не показано).
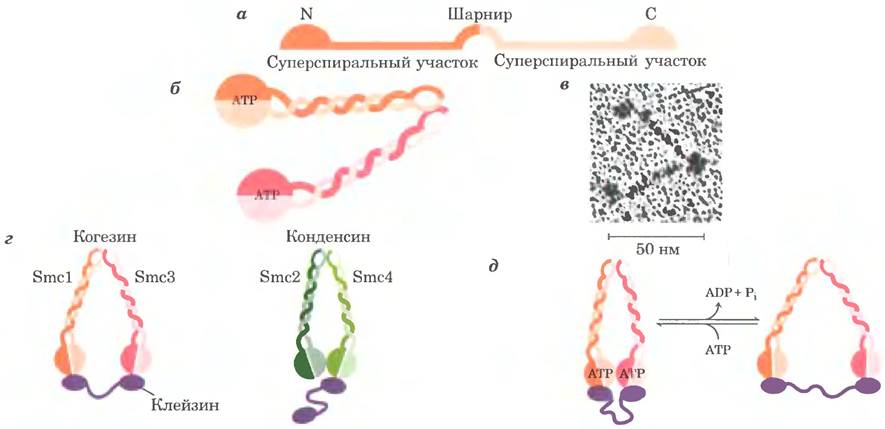
Белки семейства SMС обнаружены во всех организмах — от бактерии до человека. Эукариоты имеют два основных типа SMC-бслков — когезины и конденсины, причем оба типа этих белков связываются с регуляторными и вспомогательными белками (рис. 24-34, г). Когезины играют существенную роль в связывании между собой сестринских хроматид сразу после репликации и удерживании их в таком состоянии в процессе конденсации хромосом при переходе к стадии метафазы. Связывание важно для правильного расхождения хромосом при клеточном делении. Когезины вместе с белком третьего типа клейзином, по-видимому, образуют вокруг реплицирующихся хромосом кольцо, которое удерживает хромосомы вместе до того момента, когда в ходе клеточного деления начнется расхождение хромосом. Это кольцо может растягиваться и сжиматься в ответ на гидролиз АТР (рис. 24-34, д). Конденсины важны для конденсации хромосом при переходе клетки к митозу. В лабораторных условиях конденсины связываются с ДНК таким образом, что образуются положительные сверхвитки — связывание конденсина вызывает переход ДНК в сверхспирализованное состояние, в отличие от состояния частичного раскручивания, вызываемого связыванием нуклеосом. Пока еще не ясно, как это помогает компактной укладке хроматина, однако закручивание ДНК вокруг конденсина может играть определенную роль в конденсации ДНК. Когезины и конденсины опосредуют многие изменения в структуре хромосом в клеточном цикле эукариот (рис. 24-35).
Рис. 24-35. Моделирование роли когезинов и конденсинов в клеточном цикле эукариот. Когезины присоединяются к хромосомам в фазе G1 и удерживают вместе сестринские хроматиды в процессе репликации. При митозе с хроматидами связываются конденсины и удерживают их в конденсированном состоянии. В ходе анафазы связи с когезинами удаляются под действием так называемой сепаразной активности. После разделения хроматид конденсины начинают отделяться, и дочерние хромосомы возвращаются в неконденсированное состояние.
Бактериальная ДНК тоже сложно организована
Теперь вкратце обсудим строение бактериальных хромосом. Бактериальная ДНК упакована в структуру, называемую нуклеоидом; нуклеоид может занимать значительную часть объема клетки (рис. 24-36). Установлено, что ДНК в одной или нескольких точках прикрепляется к внутренней поверхности плазматической мембраны. О структуре нуклеоида известно гораздо меньше, чем о структуре хроматина эукариот, но постепенно проясняются детали его сложного строения. В клетках Е. coli существует своеобразный каркас, позволяющий организовать кольцевую хромосому в виде примерно 500 петлевых доменов, каждый в среднем по 10 000 п. н. (рис. 24-37), как описано выше для хроматина. Эти домены топологически ограничены; например, если происходит разрыв ДНК в одном домене, только в этом домене она и окажется релаксирована. Домены не имеют фиксированных концов. Напротив, в ходе репликации их границы постоянно смещаются по последовательности ДНК. Бактериальная ДНК, по-видимому, не имеет структуры, сопоставимой с нуклеосомами эукариот. В клетках Е. coli содержится много гистоноподобных белков (наиболее хорошо изучен состоящий из двух субъединиц белок HU Мr 19 000), но эти белки связываются и диссоциируют в течение нескольких минут, а регулярных стабильных ДНК-гистоновых комплексов обнаружено не было. Динамичные изменения в структуре бактериальной хромосомы могут быть связаны с потребностью в более быстром
доступе к генетической информации. Клеточный цикл бактериальной клетки может занимать всего 15 мин, в то время как типичная эукариотическая клетка не делится несколько часов или даже месяцев. Кроме того, у прокариот в кодировании РНК и/или белков задействована гораздо большая часть ДНК. Более интенсивные темпы клеточного метаболизма у бактерий означают, что за один и тот же отрезок времени в клетках бактерий транскрибируется или реплицируется значительно большая часть ДНК, чем у большинства эукариотических клеток.
Мы рассмотрели сложное строение ДНК и теперь готовы перейти к следующей главе, в которой обсуждается метаболизм ДНК.
Рис. 24-36. Нуклеоид Е. coli. Молекулы ДНК этих клеток обработаны красителем, который флуоресцирует при УФ-облучении. Светящиеся области представляют собой нуклеоиды. Обратите внимание, что ДНК в некоторых клетках реплицировалась, но клетки еще не разделились и поэтому содержат несколько нуклеоидов.

Рис. 24-37. Петли в хромосоме E. соli. Длина каждого такого домена около 10 000 п. н. Домены не статичны, а перемещаются по последовательности ДНК по мере продвижения репликации. Релаксации ДНК за границами того домена, в котором произошел разрыв нити, препятствуют находящиеся на границах домена элементы неизвестного состава. Эти граничные комплексы показаны в виде серых овалов. Стрелки указывают перемещение ДНК через граничные комплексы.
Краткое содержание раздела 24.3 Структура хромосом
■ Основная единица организации хроматина в клетках эукариот — нуклеосома, которая состоит из гистонов и сегмента ДНК длиной 200 п. н. Белковое ядро нуклеосомы, содержащее восемь гистонов (по две копии каждого из гистонов Н2А, H2В, Н3 и Н4), обернуто сегментом ДНК (примерно 146 п. н.) в форме левозакрученной соленоидной сверхспирали.
■ Нуклеосомы образуют 30-нм фибриллы, которые упакованы таким образом, что достигается 10 000-кратная компактизация ДНК, необходимая для размещения обычной эукариотической хромосомы в клеточном ядре. Следующие уровни организации основаны на присоединении к хромосомному остову, состоящему из гистона Н1, топоизомеразы II и SMC-белков. Белки SMC, главным образом когезины и конденсины, играют важную роль в поддержании структуры хромосом на каждой стадии клеточного цикла.
■ Бактериальные хромосомы в нуклеоиде тоже находятся в компактной упаковке, но хромосома прокариот имеет гораздо более динамичную и нерегулярную структуру, чем хроматин эукариот, что связано с краткостью клеточного цикла и высокой интенсивностью метаболизма бактерий.