Биохимия человека Том 1 - Марри Р. 1993
Биоэнергетика и метаболизм углеводов и липидов
Глюконеогенез и пентозофосфатный путь
Глюконеогенез - Метаболические пути, участвующие в глюконеогенезе (рис. 20.1)
Эти пути являются модификациями путей гликолиза и цикла лимонной кислоты. Кребс отметил, что простому обращению гликолиза препятствуют энергетические барьеры на ряде стадий; 1) между пируватом и фосфоенолпируватом, 2) между фруктозо-1,6-бисфосфатом и фруктозо-6-фосфатом, 3) между глюкозо-6-фосфатом и глюкозой, а также 4) между глюкозо-1-фосфатом и гликогеном. Эти барьеры обходятся с помощью специальных реакций.
1) В митохондриях имеется фермент пируваткарбоксилаза, который при участии АТР, биотина (витамина группы В) и СО2 превращает пируват в оксалоацетат. Функция биотина заключается в присоединении СО2 (из бикарбоната) к ферменту, далее СО2 переносится на пируват (см. ниже). Во внемитохондриальной среде клетки имеется второй фермент — фосфоенолпируваткарбоксикиназа, который катализирует превращение оксалоацетата в фосфоенолпируват. Для этой реакции требуется высокоэнергетический фосфат в форме GTP или ІТР; в результате реакции освобождается СО2. Таким образом, с помощью этих двух ферментов и лактатдегидрогеназы лактат может превращаться в фосфоенолпируват.
Существенное препятствие, однако, заключается в том, что выход оксалоацетата из митохондрии весьма затруднен. Оно преодолевается следующим образом: оксалоацетат превращается в соединение, легко диффундирующее из митохондрии во внемитохондриальный компартмент клетки, где это соединение снова превращается в оксалоацетат. Таким соединением служит малат; его образование из оксалоацетата внутри митохондрий и превращение обратно в оксалоацетат вне митохондрий катализируются малатдегидрогеназой.
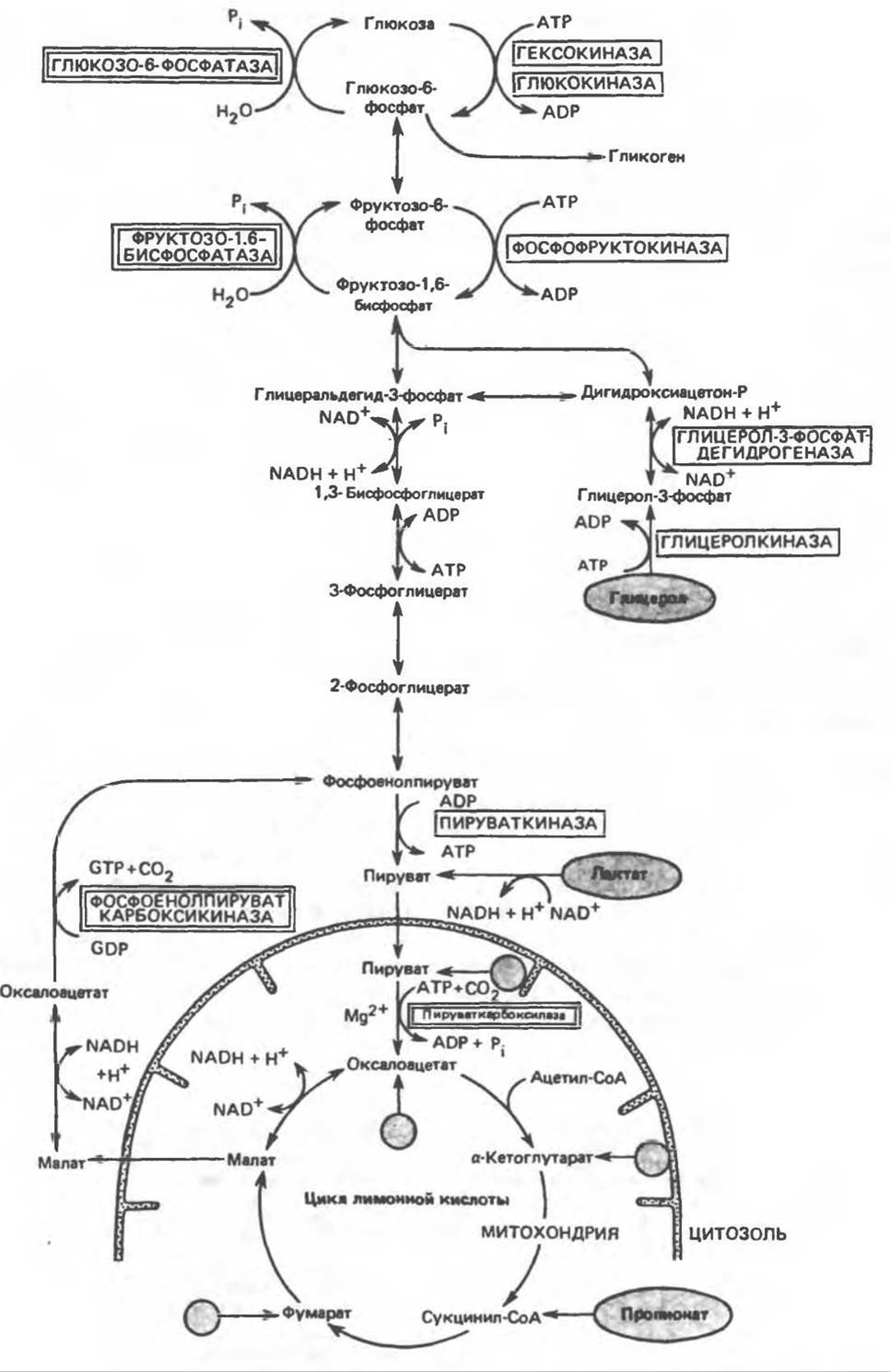
Рис. 20.1. Главные пути глюконеогенеза в печени. Кружок со стрелкой показывает места вступления в цикл глюкогенных аминокислот после переаминирования (см. также рис. 17.7). Названия ключевых ферментов глюконсогенеза заключены в двойной прямоугольник. АТР, необходимый для поддержания глюконеогенеза, генерируется в ходе окисления ацетил-СоА, который в свою очередь образуется из длинноцепочечных жирных кислот или из лактата (через пируват при действии пируватдегидрогеназы). У жвачных животных важным источником ацетил-СоА служит пропионат.
2) Превращение фруктозо-1,6-бисфосфата во фруктозо-6-фосфат, необходимое для обращения гликолиза на рассматриваемой стадии, катализируется специфическим ферментом фруктозо-1,6-бисфосфатазой. Это — ключевой фермент в том смысле, что именно его присутствием определяется, способна ли ткань ресинтезировать гликоген из пирувата и триозофосфатов. Этот фермент имеется в печени и почках, он был также обнаружен в поперечнополосатых мышцах. Считают, что в сердечной мышце и гладких мышцах он отсутствует.
3) Превращение глюкозо-6-фосфата в глюкозу катализируется другой специфической фосфатазой — глюкозо-6-фосфатазой. Она присутствует в печени и почках, но отсутствует в мышцах и жировой ткани. Наличие этого фермента позволяет ткани поставлять глюкозу в кровь.
4) Распад гликогена с образованием глюкозо-1-фосфата осуществляется фосфорилазой. Синтез гликогена идет по совершенно другому пути, через образование уридиндифосфатглюкозы, и катализируется гликогенсинтазой (см. рис. 19.1).
Взаимоотношения между этими ключевыми ферментами глюконеогенеза и гликолизом показаны на рис. 20.1. После переаминирования или дезаминирования глюкогенные аминокислоты образуют либо пируват, либо интермедиаты цикла лимонной кислоты. Поэтому описанные выше реакции могут обеспечить превращение как глюкогенных аминокислот, так и лактата в глюкозу и гликоген. Так, например, лактат превращается в пируват, который далее поступает в митохондрии, где превращается в оксалоацетат, а затем по рассмотренному выше пути — в глюкозу.
Пропионат, главный источник глюкозы у жвачных животных, вступает на путь глюконеогенеза через цикл лимонной кислоты после превращения в сукцинил-СоА. Сначала пропионат активируется при взаимодействии с АТР и СоА с помощью соответствующей ацил-СоА-синтазы. Продукт этой реакции, пропионил-СоА, присоединяет СО2 в реакции, катализируемой пропионил-СоА-карбоксилазой и превращается в D-метилмалонил-СоА (рис. 20.2). Эта реакция аналогична реакции присоединения СО2 к ацетил-СоА. катализируемой ацетил-СоА-карбоксилазой (гл. 23), в том отношении, что продуктом ее является малонильное производное и в качестве кофермента требуется биотин. D- Метилмалонил-СоА сначала превращается под действием метилмалонил-СоА-рацемазы в свой стереоизомер L-метилмалонил-СоА, а затем последний изомеризуется в сукцинил-СоА при участии метилмалонил-СоА-изомеразы, исрользующей в качестве кофермента витамин В12. Недостаток витамина В12 в организме человека и животных приводит к экскреции больших количеств метилмалоната (метилмалоновая ацидурия).
Хотя превращение в сукцинат является главным путем метаболизма пропионата, последний может быть также использован в качестве исходной молекулы для синтеза в жировой ткани и молочной железе жирных кислот с нечетным числом атомов углерода; С15- и С17-жирные кислоты обнаруживаются главным образом в липидах жвачных животных.
Глицерол является продуктом метаболизма жировой ткани; утилизировать его способны только те ткани, в которых имеется активирующий фермент глицеролкиназа. Этот фермент (ATP-зависимый) обнаружен в печени, почках и ряде других тканей. Глицеролкиназа катализирует превращение глицерола в глицерол-3-фосфат. Этот путь выходит на триозофосфатные стадии гликолиза, поскольку глицерол-3-фосфат может быть окислен NAD+ до дигидроксиацетонфосфата в присутствии глицерол-3-фосфатдегидрогеназы. Таким образом; печень и почки способны превращать глицерол в глюкозу, поступающую в кровь; при этом наряду с упомянутыми выше ферментами используются ряд ферментов гликолиза и специфические ферменты глюконеогенеза — фруктозо-1,6-бисфосфатаза и глюкозо-6-фосфатаза (рис. 20.1).
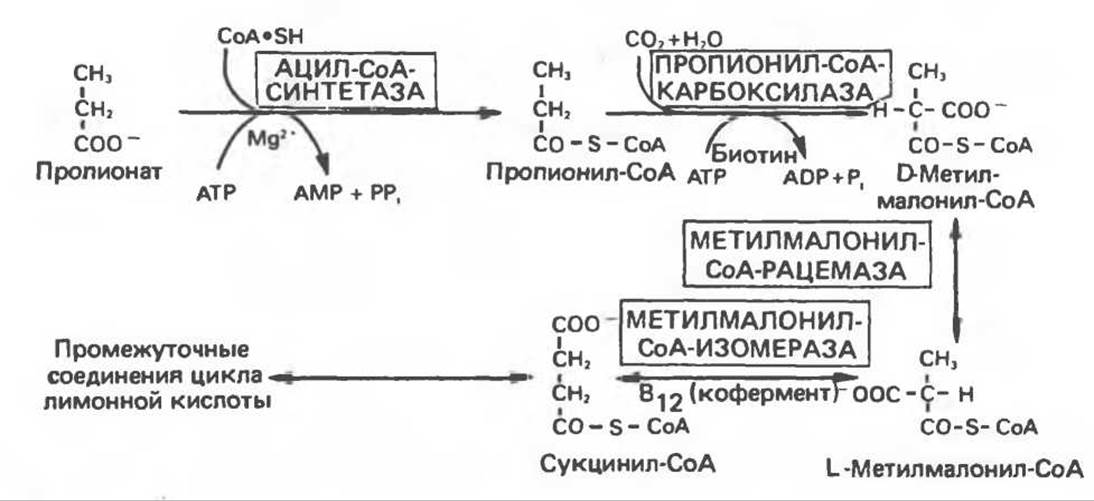
Рис. 20.2. Метаболизм пропионата.
Биотин
Биотин — один из водорастворимых витаминов группы В. Он является производным имидазола и широко распространен в натуральных пищевых продуктах (рис. 20.3). Значительную долю потребностей человека в биотине, вероятно, обеспечивают бактерии кишечника.
Биотин функционирует как компонент специфических мультисубъединичных ферментов (табл. 20.1), катализирующих реакции карбоксилирования. Он связан с апоферментом амидной связью, образуемой с ε-аминогруппой остатка лизина.
На первой стадии реакции, катализируемой пируваткарбоксилазой, карбоксилатный ион связывается с атомом N1 биотина, в результате образуется активированный интермедиат карбоксибиотин-фермент (рис. 20.4). Для протекания этой стадии необходимы НСО-3, ATP, Mg2+ и ацетил-СоА (последний в качестве аллостерического эффектора). Далее активированная карбоксильная группа переносится с интермедиата (карбоксибиотин-фермента) на пируват, при этом образуются оксалоацетат и биотинсодержащий холофермент. Длинная гибкая «ручка» (цепочка атомов) между биотином и ферментом, по-видимому, позволяет простетической группе (биотину) перемещаться от одного активного центра мультисубъединичного фермента к другому (например, с фосфокарбонатобразующего центра на пируват-связывающий центр).
Для всех апоферментов карбоксилаз имеется, по-видимому, единственный фермент, катализирующий присоединение биотина к специфическому остатку лизина. Этот фермент был назван синтетазой холокарбоксилазы. При отсутствии фермента субстраты биотинзависимых карбоксилаз накапливаются и могут быть обнаружены в моче. К числу этих метаболитов относятся лактат, ß-метилкротонат, ß-гидроксиизовалерат и ß-гидроксипропионат. У детей с недостаточностью этого фермента развивается дерматит, замедлен рост, наблюдаются алопеция, расстройство мышечной деятельности и, в некоторых случаях, заболевания, связанные с ослаблением функции иммунной системы.
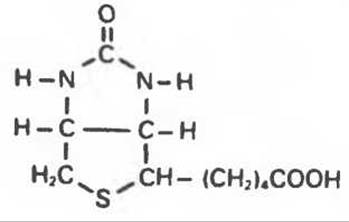
Рис. 20.3. Биотин.
Таблица 20.1. Биотинзависимые ферменты животных
|
Фермент |
Функция |
|
Пируваткарбоксилаза |
Катализирует первую реакцию пути, ведущего к превращению трехуглеродных предшественников в глюкозу (глюконеогенез) |
|
Образование оксалоацетата, участвующего в цикле лимонной кислоты |
|
|
Ацетил-СоА-карбоксилаза |
Катализирует образование малонил-СоА, поставляющего ацетатные единицы для синтеза жирных кислот |
|
Пропионил-СоА-карбоксилаза |
Превращение пропионата в сукцинат; который далее может вступать в цикл лимонной кислоты |
|
ß-Метилкротонил-СоА-карбоксилаза |
Катаболизм лейцина и некоторых изопреноидных соединений |