Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Гормоны поджелудочной железы
Инсулин
История вопроса
Островки в поджелудочной железе были обнаружены в 1860 г. Лангерганс, которому принадлежит это открытие, не представлял себе, какова их функция; не знали этого ни Меринг, ни Минковский, установившие в 1889 г., что удаление поджелудочной железы приводит к сахарному диабету. Предположение о наличии тесной связи между островками и диабетом высказали де Мейер в 1909 г. и Шарпей-Шаффер в 1917 г., но только в 1921 г. Бантинг и Бест доказали это. Экстрагировав подкисленным этанолом ткань поджелудочной железы, они выделили некий фактор, обладающий мощным гипогликемизирующим действием. Этот фактор был назван инсулином. Вскоре было установлено, что инсулин, содержащийся в островках поджелудочной железы крупного рогатого скота и свиней, активен и у человека. Не прошло и года, как этот препарат стал широко и успешно применяться для лечения диабета.
Бычий и свиной инсулин можно легко получать в больших количествах, что является важнейшим условием для успешного биохимического исследования. Именно инсулин оказался первым белком с доказанной гормональной активностью, первым белком, полученным в кристаллическом виде (Abel, 1926), первым белком, у которого была установлена аминокислотная последовательность (Sаnger et al, 1955), первым белком, синтезированным химическими методами (Du et al.; Zahn; Katsoyanis, 1964). Именно для инсулина впервые было показано, что молекула может синтезироваться в виде более крупного предшественника (Steiner et al., 1967). Кроме того, инсулин оказался первым белком, полученным для коммерческих целей с использованием технологии рекомбинантных ДНК. Но, несмотря на эти впечатляющие «первенства», механизм действия инсулина на молекулярном уровне изучен хуже, чем для большинства других гормонов.
Химические свойства
Молекула инсулина — полипептид, состоящий из двух цепей, А и В, связанных между собой двумя дисульфидными мостиками, соединяющими остаток А7 с остатком В7 и остаток А20 с остатком В19. Третий дисульфидный мостик связывает между собой остатки 6 и 11 A-цепи. Локализация всех трех дисульфидных мостиков постоянна, а А- и В-цепи у представителей большинства видов имеют по 21 и 30 аминокислот соответственно. Ковалентная структура человеческого инсулина (мол. масса 5734) показана на рис. 51.1, сведения об аминокислотных заменах в инсулинах различных видов содержатся в табл. 51.2. В обеих цепях во многих положениях встречаются замены, не оказывающие влияния на биологическую активность гормона, однако наиболее часты замены по положениям 8,9 и 10 A-цепи. Из этого следует, что данный участок молекулы не имеет критического значения для биологической активности инсулина. Однако некоторые участки и области молекулы инсулина обладают высокой консервативностью.

Рис. 51.1. Ковалентная структура инсулина человека. (Reproduced, with permission, from Ganong W.F. Review of Medical Physiology, 13th ed., Appleton and Lange, 1987.)
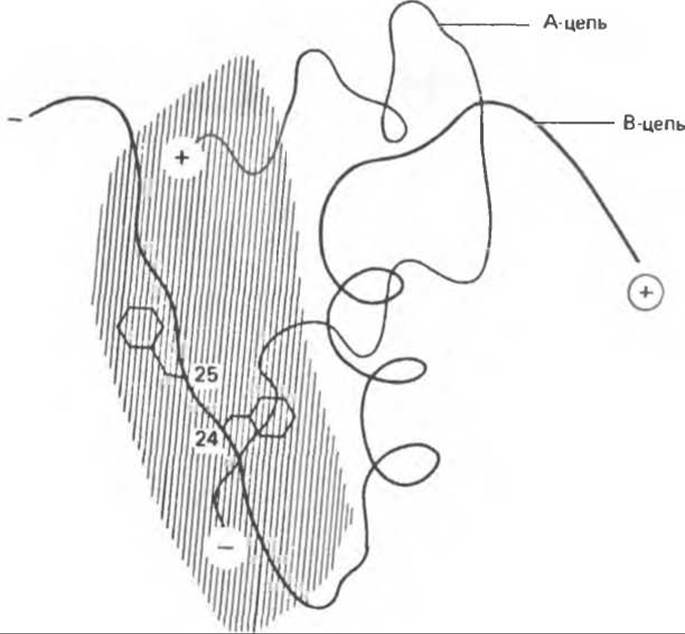
Рис. 51.2. Участок молекулы инсулина, отвечающий за его биологическую активность. Данная схема молекулы инсулина построена по результатам рентгеноструктурной кристаллографии. Заштрихованная область соответствует той части инсулина, которой отводят главную роль в реализации биологической активности гормона. Остатки Phe в положениях В24 и В25 — это те сайты, мутации в которых влияют на биологическую активность инсулина. N-концы А- и В-цепей инсулина показаны знаком «+», тогда как С- концы — знаком «—». (Redrawn and reproduced, with permission, from Tager H.S. Abnormal products of the human insulin gene, Diabetes, 1984, 33, 693.)
К ним относятся 1) положения трех дисульфидных мостиков, 2) гидрофобные остатки в С-концевом участке В-цепи и 3) С- и N-концевые участки А-цепи. Использование химических модификаций и замен отдельных аминокислот шести этих участков помогли идентифицировать сложный активный центр (рис. 51.2). Расположенный на С-конце В-цепи гидрофобный участок участвует также в димеризации инсулина.
Таблица 51.2. Различия в структуре инсулина у представителей разных видов млекопитающих. (Modified and reproduced, with permission, from Banong W. F.: Review of Medical Physiology. 13th ed., Appleton and Lange, 1987.)
|
Вид |
Аминокислотозамены относительно инсулина человека |
|||
|
Положение цепи в А- |
Положение в В-цени |
|||
|
8 |
9 |
10 |
30 |
|
|
Человек |
Thr- |
Ser- |
Не |
Thr |
|
Свинья, собака, кашалот |
Thr- |
Ser- |
Ilе |
Ala |
|
Кролик |
Thr- |
Ser- |
Ikе |
Ser |
|
Крупный рогатый скот, коза |
Ala- |
Ser- |
Val |
Ala |
|
Овца |
Ala- |
Gly- |
Val |
Ala |
|
Лошадь |
Thr- |
Gly- |
Ile |
Ala |
|
Сейвал |
Ala- |
Ser- |
Thr |
Ala |
Как явствует из табл. 51.2, между инсулином человека, свиньи и быка существует большое сходство.
Инсулин свиньи отличается от человеческого инсулина одной-единственной аминокислотной заменой: вместо треонина в положении 30 В-цепи находится аланин. В бычьем инсулине помимо этого треонин А8 заменен на аланин, а изолейцин А10 — на валин. Эти замены практически не влияют на биологическую активность гормона и очень слабо влияют на его антигенные свойства. Хотя у большинства больных, получавших гетерологичный инсулин, обнаруживаются циркулирующие в небольшом титре антитела против введенного гормона, некоторые пациенты демонстрируют титр антител клинически значимой величины. До тех пор пока человеческий инсулин не научились получать с помощью методов генной инженерии, для терапевтических целей использовали обычно бычий и свиной инсулины. Несмотря на значительные различия в первичной структуре, все три инсулина имеют сходную биологическую активность (25—30 МЕд/мг сухого веса).
Инсулин образует очень интересные сложные структуры. Цинк, концентрация которого в B-клетках достигает высоких значений, формирует комплексы с инсулином и проинсулином. Инсулины всех позвоночных образуют изологичные димеры с помощью водородных связей между пептидными группами остатков В24 и В26 двух мономеров, которые при высоких концентрациях в свою очередь реорганизуются в гексамеры, содержащие по два атома цинка каждый. Наличие такой высоко упорядоченной структуры облегчило изучение кристаллической структуры инсулина. При физиологических концентрациях инсулин находится, вероятно, в мономерной форме.
Биосинтез
А. Предшественники инсулина. Инсулин синтезируется в виде препрогормона (мол. масса 11 500). Он может служить примером пептида, образующегося в результате различных преобразований из более крупной молекулы предшественника. Последовательность и субклеточная локализация соответствующих биохимических превращений показаны на рис. 51.3. Состоящая из 23 аминокислот гидрофобная лидерная последовательность (пре-фрагмент) направляет молекулу-предшественник в цистерну эндоплазматического ретикулума и там отделяется. В результате образуется молекула проинсулина (мол. масса 9000), принимающая конформацию, необходимую для образования нужных дисульфидных мостиков. Как показано на рис. 51.4, молекула проинсулина имеет следующее строение, считая от аминоконца:
![]()
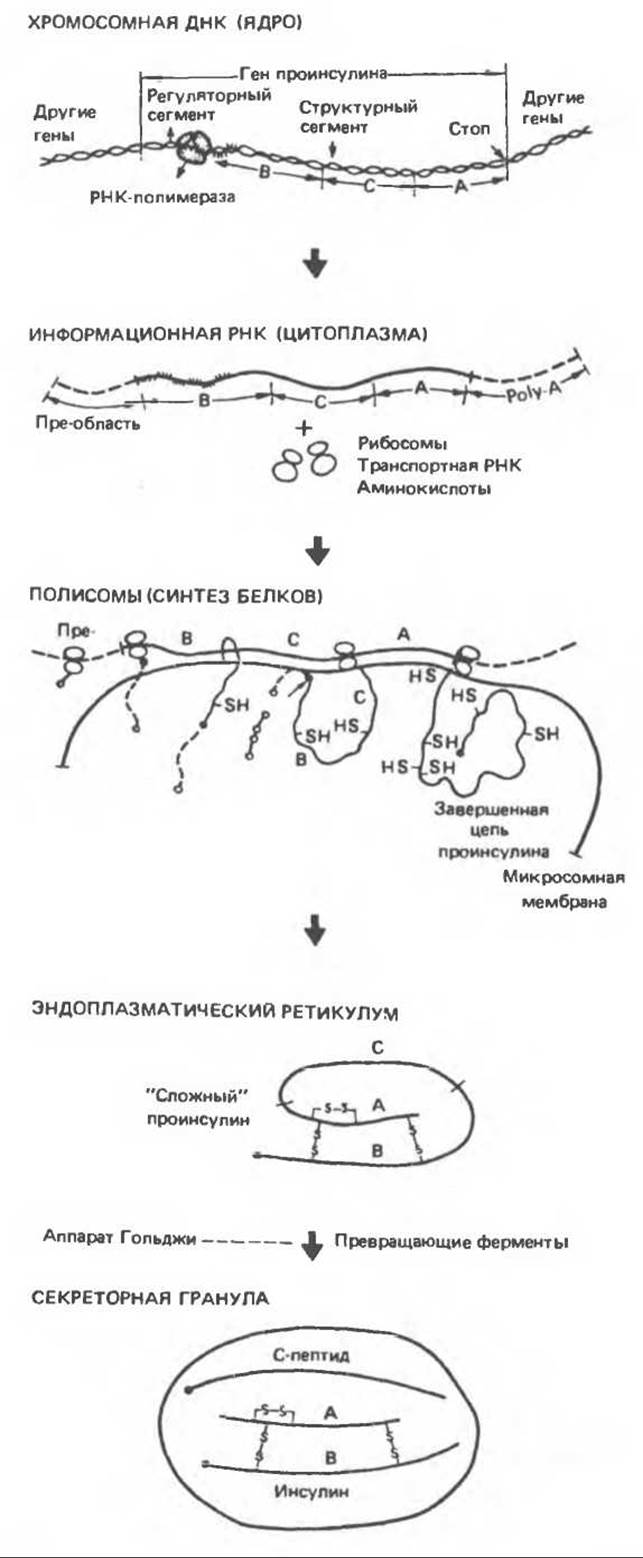
Рис. 51.3. Биосинтез инсулина с образованием короткоживущего предшественника. Буквами А, В и С обозначены А и В-цепи инсулина и связующий (С) пептид. Лидерная последовательность из 23 аминокислот, закодированная в сегменте мРНК, расположенном рядом с тем сегментом, который детерминирует В-цепь (прерывистая линия), после образования отщепляется, вероятно, еще до завершения синтеза остальной части молекулы проинсулина. (Reproduced, with permission, from Steiner D. F., Erros in insulin biosynthesis, N. Engl. J. Med., 1976, 294, 952.)
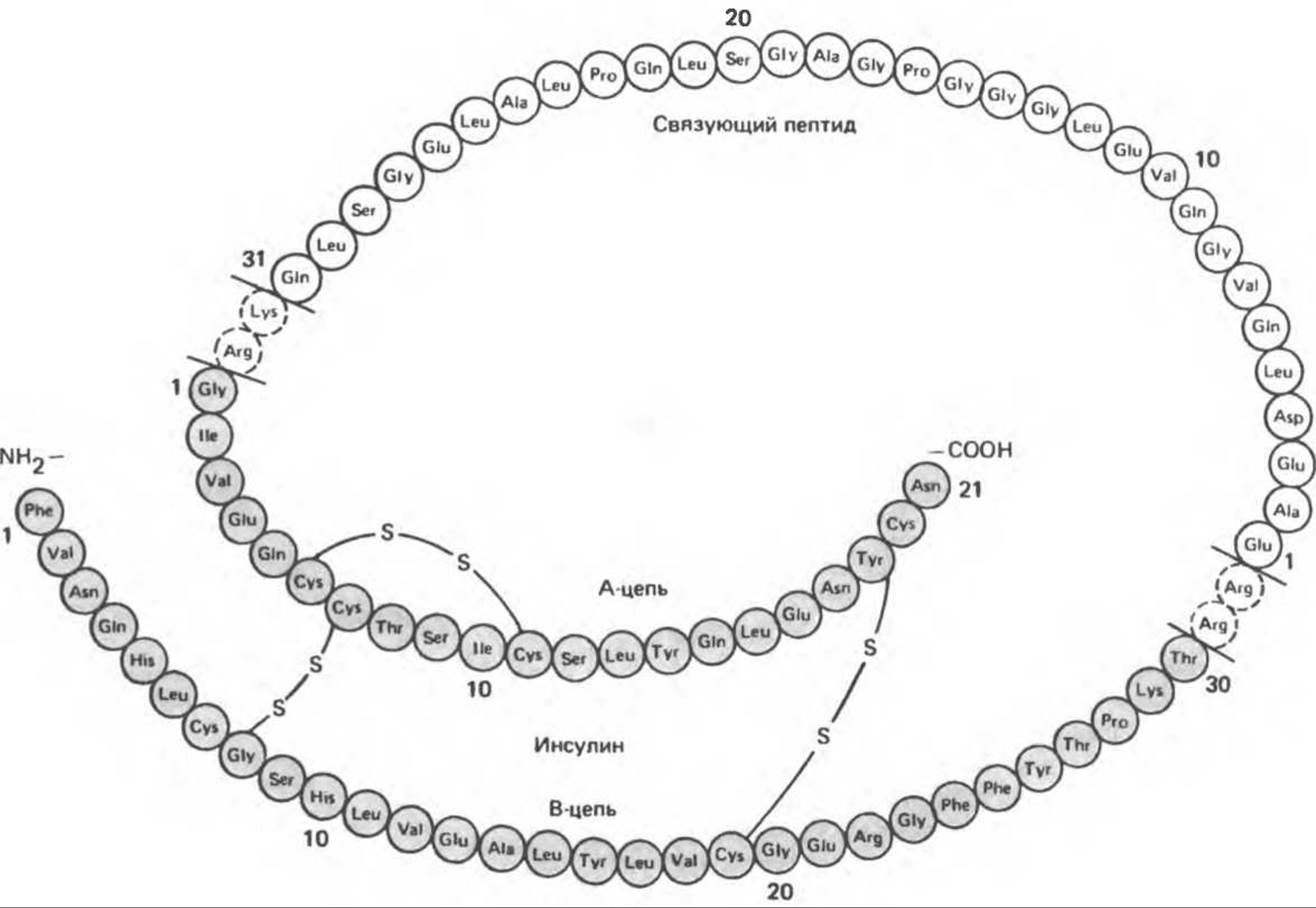
Рис. 51.4. Структура проинсулина человека. Молекулы инсулина и С-пептида связаны между собой с помощью двух дипептидных линкеров, расположенных по обе стороны от С-пептида. (Slightly modified and reproduced, with permission, from Karam J. H., Sabler P. R., Forsham P. H. Pancreatic hormones and diabetes mellitus. In: Basic and Clinical Endocrinology, 2nd ed., Greenspan F. S., Forsham P. H.(eds.), Appleton and Lange, 1986.)
Молекула проинсулина расщепляется в нескольких специфических участках с образованием эквимолярных количеств зрелого инсулина и С-пептида. Эти ферментативные превращения, показанные схематически на рис. 51.5, начинаются с протеиназы, обладающей трипсиноподобной активностью — ферментом, отщепляющим с каждой из сторон С- пептида по две основные аминокислоты: дипептид Arg31-Arg32 на N-конце С-пептида и дипептид Lys64-Arg65 — на С-конце С-пептида2.
1 С-пептид от англ. connecting—связующий. — Прим, перев.
2 Указанное положение дипептидов соответствует их положению в полной молекуле проинсулина при отсчете с N-конца. — Прим. перев.
Б. Предшественники других гормонов островковых клеток. Синтез других гормонов островковых клеток также требует ферментативного превращения молекул-предшественников с большей молекулярной массой. Строение молекул панкреатического полипептида, глюкагона и соматостатина в сравнении со строением инсулина схематически показано на рис. 51.6. В образовании этих гормонов участвуют различные комбинации эндопротеолитических (трипсиноподобных) и экзопротеолитических (подобных карбоксипептидазе-В) ферментов, поскольку обладающие гормональной активностью последовательности могут обнаруживаться в различных участках молекулы-предшественника: соматостатин — на карбоксильном конце молекулы, панкреатический полипетид — на аминоконце, инсулин — на обоих концах и глюкагон — в средней части.
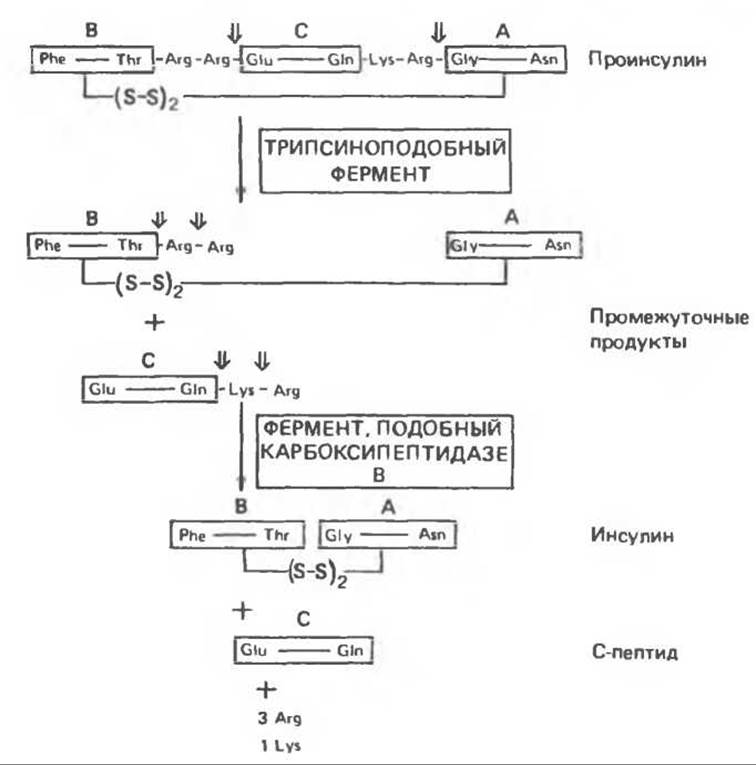
Рис. 51.5. Стадии расщепления проинсулина человека при совместном действии протеиназ, подобных трипсину, и карбоксипептидазе В. Стрелками показано, в каких местах происходит расщепление молекулы. (Redrawn and reproduced, with permission, from Steiner D. F., Tager H. S. p. 927. In: Endocrinology, Vol. 2., DeGroot L. J. (ed.), Grune and Stratton, 1979.)
В. Субклеточная локализация синтеза инсулина и формирование гранул. Синтез инсулина и его упаковка в гранулы происходит в определенном порядке (рис. 51.7). Проинсулин синтезируется на рибосомах шероховатого эндоплазматического ретикулума. Затем в цистернах этой органеллы происходит ферментативное отщепление лидерной последовательности (пре-сегмент), образование дисульфидных мостиков и складывание молекулы (рис. 51.3). После этого молекула проинсулина переносится в аппарат Гольджи, где начинаются протеолиз и упаковка в секреторные гранулы. Созревание гранул продолжается по мере продвижения по цитоплазме в направлении плазматической мембраны. Как проинсулин, так и инсулин соединяются с цинком, образуя гексамеры, но поскольку около 95% проинсулина превращается в инсулин, то именно кристаллы последнего придают гранулам их морфологические особенности. Наряду с инсулином в гранулах содержатся также эквимолярные количества С-пептида, однако эти молекулы не образуют кристаллических структур. При соответствующей стимуляции зрелые гранулы сливаются с плазматической мембраной, выбрасывая свое содержимое во внеклеточную жидкость путем эмиоцитоза.
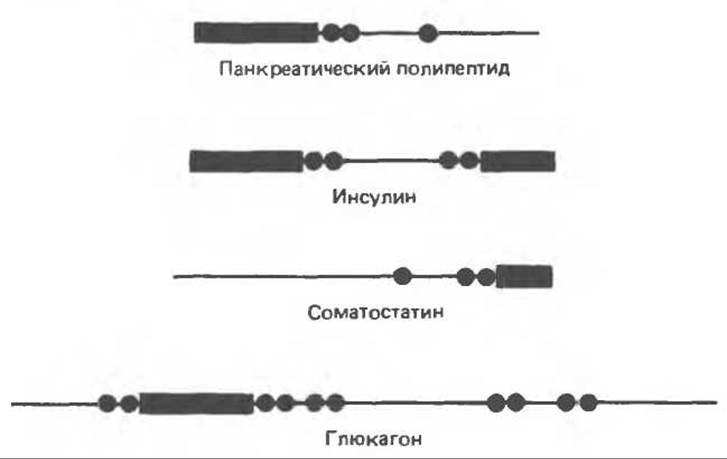
Рис. 51.6. Схема строения четырех основных продуктов эндокринных клеток поджелудочной железы. Черными полосками показана часть молекулы предшественника, соответствующая указанному в надписи гормону, тонкой линией обозначены остальные участки пептидной цепи молекулы-предшественника. Места расположения двухосновных аминокислот (аргинина или лизина), где происходит расщепление молекулы-предшественника, обозначены черными кружками. Молекула проинсулина изображена в виде линейной структуры, в которой дисульфидные связи не показаны. В действительности молекула проинсулина имеет последовательность: В-цепь — С-пептид A-цепь. (Redrawn and reproduced, with permission, from Tager H.S. Abnormal products of the human insulin gene. Diabetes. 1984. 33. 693.)
Г. Свойства проинсулина и С-пептида. Длина проинсулинов колеблется от 78 до 86 аминокислот, причем эти различия обусловлены длиной С-пептида. Проинсулин имеет ту же растворимость и изоэлектрическую точку, что и инсулин. Он также образует гексамеры с кристаллами цинка и реагирует с антисывороткой к инсулину. Биологическая активность проинсулина составляет менее 5% биологической активности инсулина. Отсюда следует, что большая часть активного центра инсулина в молекуле предшественника замаскирована. Некоторая часть проинсулина секретируется вместе с инсулином, а в определенных ситуациях (опухоль из островковых клеток) он высвобождается в больших количествах, чем в норме. Поскольку период полужизни проинсулина в плазме значительно выше, чем у инсулина, и при этом проинсулин дает сильную перекрестную реакцию с антисывороткой к инсулину, уровень «инсулина», определяемый радиоиммунологическим методом, в некоторых случаях может превышать содержание биологически активного гормона.
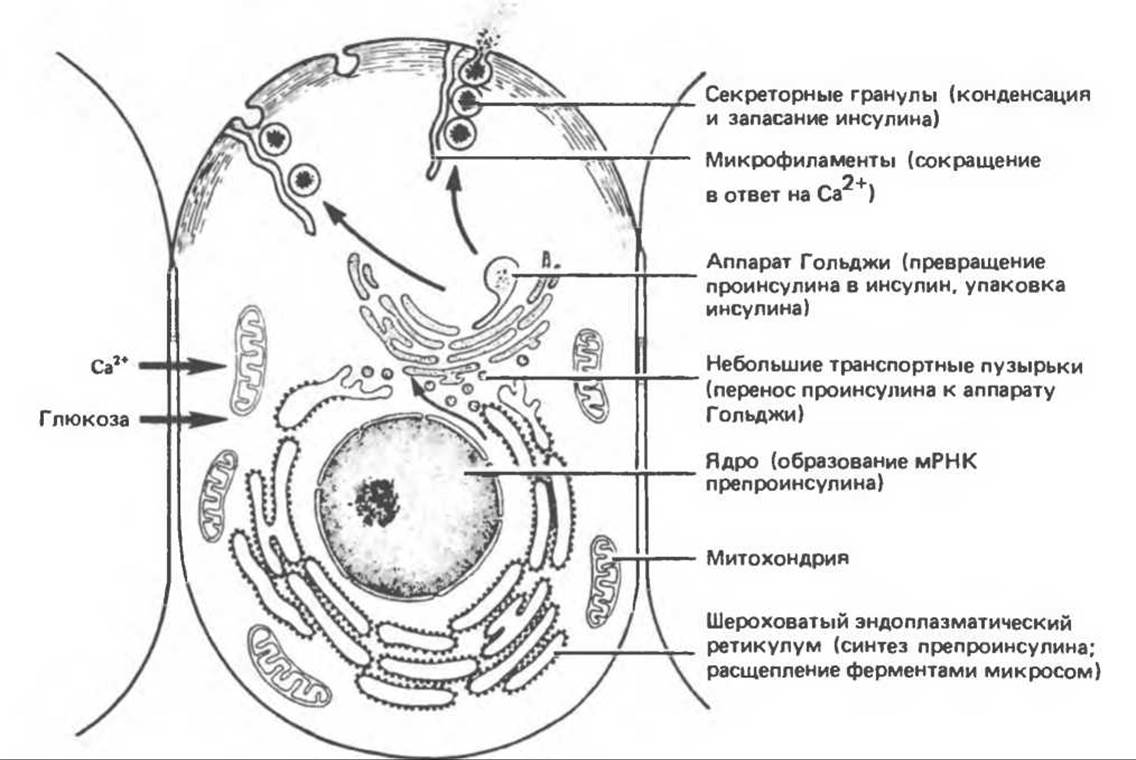
Рис. 51.7. Структурные компоненты В-клетки поджелудочной железы, участвующие в индуцированных глюкозой биосинтезе и секреции гормона. На схеме секреторные гранулы прилегают к микрофиламентам, которые сокращаются под влиянием кальция. (Based on data presented by Orci L. A portrait of the pancreatic В cell, Diabetologia, 1974, 10, 163.) (Modified and reproduced, with permission, from Junqueira L. C., Carneiro J., Long J. A., Basic Histology. 5th ed., Appleton and Lange, 1986.)
Какой-либо биологической активности С-пептида не обнаружено. Эта молекула обладает иными антигенными свойствами, чем инсулин и проинсулин, поэтому иммунологическое определение С-пептида позволяет отличить эндогенносекретируемый инсулин от вводимого гормона и дает возможность судить о количестве эндогенного инсулина в тех случаях, когда его прямое определение оказывается невозможным из-за наличия инсулиновых антител. С-пептиды представителей различных видов характеризуются высокой частотой аминокислотных замен, что подтверждает положение о вероятном отсутствии биологической активности у этого фрагмента.
Д. Предшественники пептидов, родственных инсулину. Структурная организация молекулы прогормона неспецифична для предшественника инсулина. Предшественники близкородственных инсулину пептидных гормонов (релаксина и инсулиноподобных факторов роста) имеют такую же организацию (рис. 51.8). У всех этих гормонов последовательности А- и В-цепей в молекуле предшественника имеют на карбоксильных и аминоконцах высокогомологичные участки, соединяющиеся между собой связующим пептидом. В пептидных предшественниках инсулина и релаксина по обе стороны от связующего пептида расположены по две основные аминокислоты, соединяющие его с А- и В-цепями. После возникновения между А- и В-цепями дисульфидных мостиков связующий пептид вырезается в результате эндопротеолиза, и молекула превращается в пептидный гормон, состоящий из двух (А и В) цепей. Инсулиноподобные факторы роста, будучи высокогомологичными инсулину и релаксину по своей первичной структуре, тем не менее имеют одно важное отличие: в молекуле их предшественника отсутствуют участки, по которым происходит отщепление связующего пептида, и поэтому активные гормоны сохраняют структуру единой полипептидной цепи.
Е. Ген инсулина человека. Ген человеческого инсулина (рис. 51.9) локализован в коротком плече хромосомы 11. У большинства млекопитающих экспрессируется один ген инсулина, организованный подобно человеческому гену, но у крыс и мышей имеются два неаллельных гена. В каждом из них закодирован особый проинсулин, дающий начало двум различным активным молекулам инсулина. В настоящее время разработан метод получения человеческого инсулина в бактериальных экспрессирующих системах с использованием технологии рекомбинантных ДНК. Таким образом, проблему получения этого гормона в количествах, необходимых для больных диабетом, можно считать решенной.
Ж. Аномальные продукты гена инсулина человека. Знание структуры инсулинового гена и инсулиновой молекулы позволяет выявлять аномальные продукты гена, что в свою очередь дает дополнительную информацию о функции данного гормона. Выявлены три мутации этого гена, причем для каждой из них идентифицирована молекулярная основа дефекта. В одном случае в результате мутации единичного основания на месте фенилаланина-В24 оказался серин, в другом (опять-таки в результате единичной мутации) произошла замена фенилаланина-В25 на лейцин. В третьем случае изменился процессинг проинсулина в активный гормон: мутация нарушила отщепление 3'-конца С-пептида на границе с А-цепью. В основе этого дефекта лежит замена дипептида Lys-Arg в этом месте полипептидной цепи на Lys-X, в результате которой трипсиноподобное расщепление оказалось невозможным. Выявлению описанных мутаций способствовала их локализация в активном центре молекулы инсулина, в результате чего у соответствующих носителей 1) имеет место гиперинсулинемия, 2) отсутствуют признаки инсулинорезистентности, 3) снижена биологическая активность циркулирующего в крови инсулина и 4) отмечается нормальная реакция на экзогенный инсулин. По крайней мере еще четыре нуклеотидные замены были идентифицированы у «здоровых» людей. Эти мутации локализованы во вставочных (т. е. некодирующих) последовательностях, и на функциональную активность молекулы инсулина они не повлияли.
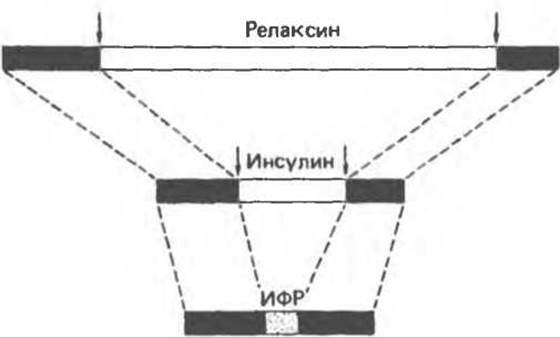
Рис. 51.8. Схематическое изображение структуры предшественников родственных инсулину пептидов. Гомологичные участки релаксина, инсулина и инсулиноподобного фактора роста изображены в виде черных полос. Аминокислотные последовагельности, соединяющие В- и А-цепи в молекуле предшественников релаксина и инсулина, обозначены светлыми полосами. В ходе процессинга предшественников с образованием соответствующих продуктов, состоящих из двух цепей, эти связующие последовательности удаляются (вертикальные стрелки). Аминокислотная последовательность инсулиноподобного фактора роста, соответствующая таким связующим пептидам, но не удаляемая в ходе процессинга, изображена участком с точками. Инсулиноподобный фактор роста состоит лишь из одной пептидной цепи. (Redrawn and reproduced, with permission. from Tager H. S. Abnormal products of the human insulin gene, Diabetes, 1984, 33, 693.)
![]()
Рис. 51.9. Схематическое изображение структуры гена человеческого инсулина. Области, заштрихованные диагональными линиями, соответствуют нетранслируемым областям мРНК. Светлые участки соответствуют вставочным последовательностям, участки, выделенные пунктиром, — кодирующим последовательностям. Буквами L, В, С и А обозначены последовательности, кодирующие лидерный (сигнальный) пептид, В-цепь инсулина. С-пептид и А-цепь инсулина соответственно. Следует обратить внимание на то, что кодирующая последовательность для С-пептида разделена вставочной последовательностью. Масштаб в схеме выдержан. (Redrawn and reproduced, with permission. from Tager H. S. Abnormal products of the human insulin gene. Diabetes, 1984, 33, 639.)
Регуляция секреции инсулина
Поджелудочная железа человека секретирует до 40—50 ед. инсулина в сутки, что соответствует 15—20% общего количества гормона в железе. Секреция инсулина — энергозависимый процесс, происходящий с участием системы микротрубочек и микрофиламентов островковых В-клеток и ряда медиаторов.
А. Глюкоза. Повышение концентрации глюкозы в крови — главный физиологический стимул секреции инсулина. Пороговой для секреции инсулина является концентрация глюкозы натощак 80—100 мг%, а максимальная реакция достигается при концентрации глюкозы 300—500 мг%. Секреция инсулина в ответ на повышение концентрации глюкозы носит двухфазный характер (рис. 51.10). Немедленный ответ, или первая фаза реакции, начинается в пределах 1 мин после повышения концентрации глюкозы и продолжается в течение 5—10 мин. Затем наступает более медленная и продолжительная вторая фаза, обрывающаяся сразу после удаления глюкозного стимула. Согласно существующим представлениям, наличие двух фаз ответной реакции инсулина отражает существование двух различных внутриклеточных компартментов, или пулов, инсулина. Абсолютная концентрация глюкозы в плазме — не единственная детерминанта секреции инсулина. В-клетки реагируют и на скорость изменения концентрации глюкозы в плазме.
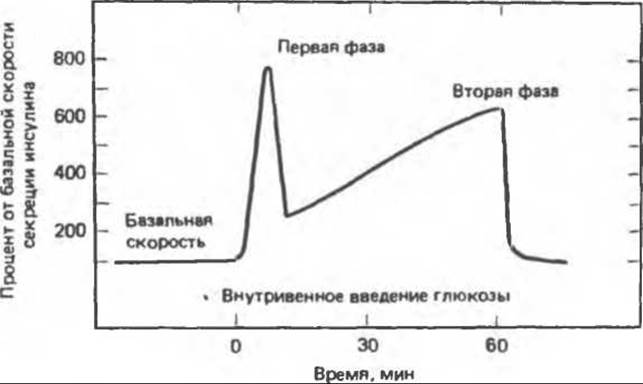
Рис. 51.10. Двухфазный характер секреции инсулина в ответ на повышение концентрации глюкозы в плазме крови.
При пероральном введении глюкозы происходит гораздо более сильная стимуляция секреции инсулина, чем при ее внутривенном введении. Отсюда следует, что на секрецию инсулина помимо глюкозы влияют также и различные гормоны желудочнокишечного тракта, такие, как секретин, холецистокинин, гастрин и энтероглюкагон. Однако наибольшая роль в этом процессе принадлежит желудочному ингибиторному полипептиду (ЖИП).
Предполагаются два разных механизма регуляции глюкозой секреции инсулина. Согласно одной гипотезе, глюкоза взаимодействует с рецептором, локализованным, вероятно, на поверхностной мембране В-клетки, что и приводит к активации механизма секреции. Вторая гипотеза исходит из того, что в стимуляции секреции инсулина участвуют внутриклеточные метаболиты или скорость таких метаболических путей, как пентозофосфатный шунт, цикл лимонной кислоты или гликолиз. Обе гипотезы нашли экспериментальные подтверждения.
Б. Гормональные факторы. На высвобождение инсулина влияет множество гормонов. а-Адренергические агонисты, особенно адреналин, подавляют секрецию инсулина даже при стимуляции этого процесса глюкозой. ß-Адренергические агонисты стимулируют секрецию инсулина, вероятно, путем повышения концентрации внутриклеточного сАМР (см. ниже). Именно этот механизм, по-видимому, лежит в основе действия желудочного ингибиторного пептида, который повышает секрецию инсулина, а также в основе эффектов высоких концентраций ТТГ, АКТГ, гастрина, секретина, холецистокинина и энтероглюкагона.
При хроническом воздействии избыточных количеств гормона роста, кортизола, плацентарного лактогена, эстрогенов и прогестинов секреция инсулина также повышается. Поэтому и неудивительно, что на поздних сроках беременности секреция инсулина значительно возрастает.
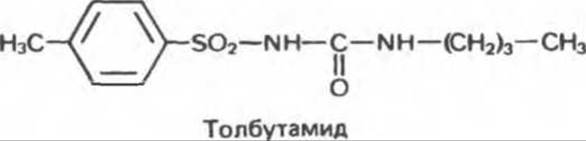
В. Фармакологические агенты. Секрецию инсулина стимулируют многие лекарственные препараты, однако в терапевтических целях чаще всего используются производные сульфонилмочевины. Для лечения диабета типа II (инсулиннезависимого) широко применяют такие средства, как толбутамид, который стимулирует секрецию инсулина иным способом, чем глюкоза.
Г. Внутриклеточные медиаторы секреции. При стимуляции секреции инсулина глюкозой возрастает потребление О, и использование АТР. Это сопряжено с индуцированной К+ деполяризацией мембраны, что приводит к быстрому проникновению в клетку Са2+ по потенциал-зависимому каналу. Слияние инсулин-содержащих секреторных гранул с плазматической мембраной и происходящая в результате секреция инсулина — процесс, зависимый от кальция. Стимуляция секреции инсулина глюкозой происходит и с участием метаболитов фосфатидилинозитола (гл. 44).
В процессе секреции инсулина участвует и сАМР, который потенциирует эффекты глюкозы и аминокислот. Этот нуклеотид может стимулировать высвобождение Са2+ из внутриклеточных органелл или активировать киназу, фосфорилирующую какой-то компонент системы микрофиламенты — микротрубочки (что обусловливает ее чувствительность к Са2+ и способность к сокращению). Замена внеклеточного Na+ на какой-либо другой одновалентный катион ослабляет эффекты глюкозы и других стимуляторов секреции инсулина; Na+, возможно, регулирует внутриклеточную концентрацию Са2+ через систему совместного транспорта.
Метаболизм инсулина
В отличие от инсулиноподобных факторов роста инсулин не имеет белка-носителя в плазме. Поэтому в норме период его полужизни не достигает и 3—5 мин. Метаболические превращения инсулина происходят в основном в печени, почках и плаценте. Около 50% этого гормона исчезает из плазмы за один пассаж через печень. В метаболизме инсулина участвуют две ферментные системы. Первая представляет собой инсулин-специфическую протеиназу, обнаруживаемую во многих тканях, но в наибольшей концентрации — в органах, перечисленных выше. Эта протеиназа была выделена из скелетных мышц и очищена. Установлено, что ее активность зависит от сульфгидрильных групп и проявляется при физиологических значениях pH. Вторая система — глутатион-инсулин-трансгидрогеназа. Этот фермент восстанавливает дисульфидные мостики, после чего отделенные друг от друга А- и В-цепи быстро расщепляются. Какой из двух механизмов наиболее активен в физиологических условиях, не ясно; не ясно также, является ли каждый из них регулируемым.
Физиологические эффекты инсулина
О том, сколь велика роль инсулина в углеводном, белковом и липидном обмене, яснее всего свидетельствуют последствия инсулиновой недостаточности у человека. Основным признаком сахарного диабета является гипергликемия, развивающаяся в результате 1) пониженного проникновения глюкозы в клетки, 2) снижения утилизации глюкозы различными тканями и 3) повышения образования глюкозы (глюконеогенеза) в печени. Ниже мы рассмотрим все эти процессы более подробно.
Полиурия, полидипсия и потеря веса, несмотря на адекватное потребление калорий, — таковы главные симптомы инсулиновой недостаточности. Чем они объясняются? Если в норме уровень глюкозы в плазме у человека редко превышает 120 мг%, то у больных с инсулиновой недостаточностью он, как правило, бывает значительно выше. Когда содержание глюкозы в плазме достигает определенных значений (у человека это обычно выше 180 мг%), максимальная способность реабсорбции глюкозы в почечных канальцах оказывается превышенной и сахар выделяется с мочой (глюкозурия). Объем мочи при этом увеличивается из-за осмотического диуреза, что обязательно сопровождается вначале потерей жидкости (полиурия), затем обезвоживанием организма, жаждой и чрезмерным потреблением воды (полидипсия). Глюкозурия вызывает значительную потерю калорий (4,1 ккал на каждый грамм экскретируемой глюкозы), что в сочетании с потерей мышечной и жировой ткани приводит к резкому похуданию, несмотря на повышенный аппетит (полифагия) и нормальное или увеличенное потребление калорий.
В отсутствие инсулина снижается биосинтез белка, что отчасти объясняется уменьшением транспорта аминокислот в мышцы (аминокислоты служат субстратами для глюконеогенеза). Таким образом, инсулиновая недостаточность у человека сопровождается отрицательным азотным балансом. Характерное для этой ситуации отсутствие антилиполитического действия инсулина, равно как и его липогенного действия, приводит к тому, что содержание жирных кислот в плазме возрастает. Когда оно достигает уровня, превышающего способность печени окислять жирные кислоты до СО2, в крови накапливаются ß-гидроксимасляная и ацетоуксусная кислоты (кетоз). Вначале организм компенсирует накопление этих органических кислот увеличением количества выдыхаемого СО2. Однако если развитие кетоза не сдерживается введением инсулина, то развивается тяжелый метаболический ацидоз и больной погибает от диабетической комы. Механизм инсулиновой недостаточности схематически представлен на рис. 51.11.
А. Влияние на транспорт глюкозы через мембрану.
Внутриклеточная концентрация свободной глюкозы значительно ниже ее внеклеточной концентрации. Большинство имеющихся данных свидетельствует о том, что скорость транспорта глюкозы через плазматическую мембрану мышечных и жировых клеток определяет интенсивность фосфорилирования глюкозы и ее дальнейший метаболизм. D-глюкоза и другие сахара с аналогичной конфигурацией по С1-С3 (галактоза, D-ксилоза и L-арабиноза) проникают в клетки путем облегченной диффузии, опосредованной переносчиком. Во многих клетках инсулин усиливает этот процесс (рис. 51.12), что обусловливается увеличением числа переносчиков (Vmaх-эффект), а не повышением сродства связывания (Kм-эффект). Согласно имеющимся данным, в жировых клетках это происходит путем мобилизации переносчиков глюкозы из неактивного их пула в аппарате Гольджи с дальнейшим направлением их к активному участку плазматической мембраны. Такая транслокация переносчиков — процесс, зависимый от температуры и энергии и независимый от синтеза белков (рис. 51.13).
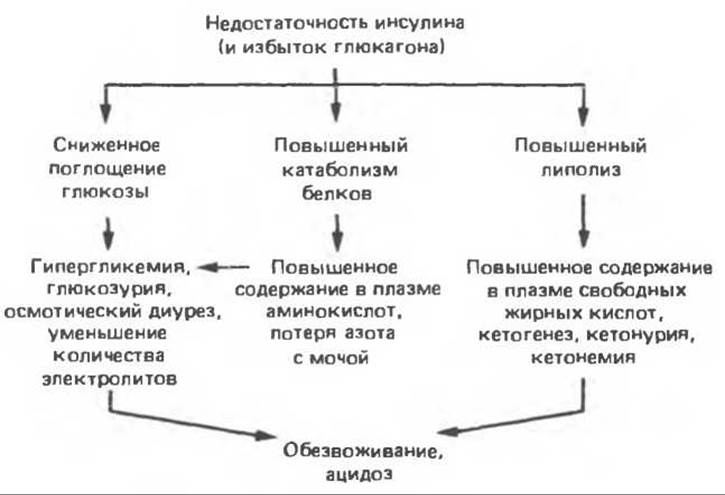
Рис. 51.11. Патофизиология инсулиновой недостаточности. (Courtesy of R. J. Havel.)
Печеночные клетки представляют собой важное исключение из этой схемы. Инсулин не стимулирует облегченной диффузии глюкозы в гепатоциты, но усиливает ее приток косвенным путем, индуцируя глюкокиназу — фермент, превращающий глюкозу в глюкозо-6-фосфат. В результате быстро протекающего фосфорилирования концентрация свободной глюкозы в гепатоцитах поддерживается на очень низком уровне, что способствует проникновению глюкозы в клетки путем простой диффузии по градиенту концентрации.
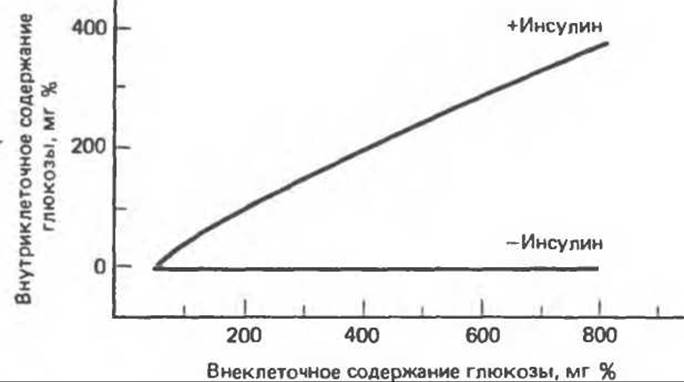
Рис. 51.12. Проникновение глюкозы в мышечные клетки.
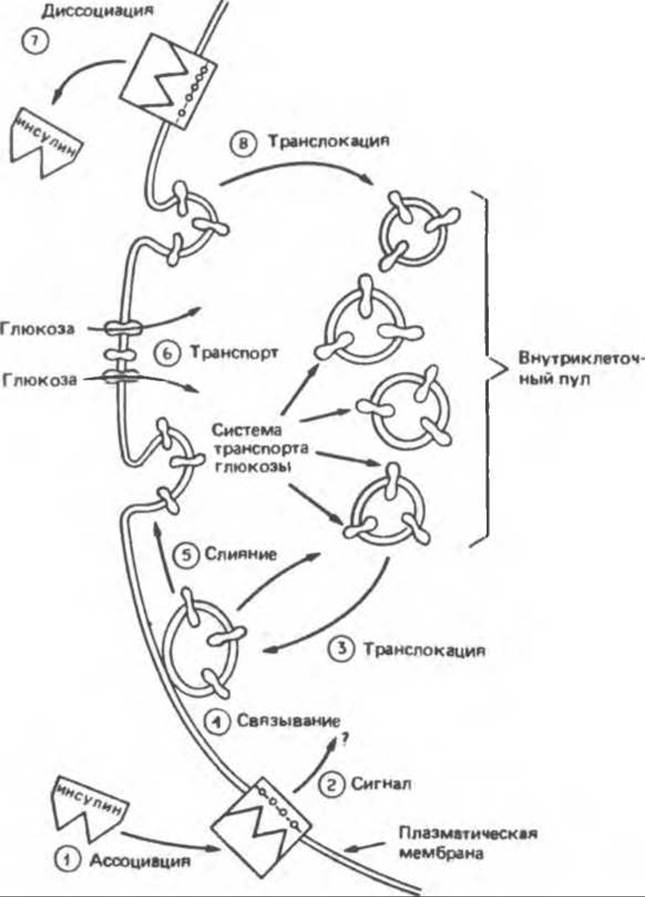
Рис. 51.13. Транслокация переносчиков глюкозы под влиянием инсулина. (Reproduced, with permission, from Karnieli E. et al. Insulin-stimulated translocation of glucose transport systems in the isolated rat adipose cell, J. Biol, Chem., 1981, 256, 4772, Courtesy of S. Cushman.)
Инсулин способствует также проникновению в клетки аминокислот (особенно в мышечные клетки) и стимулирует перемещение К+, Са2+, нуклеозидов и органического фосфата. Эти эффекты не зависят от влияния инсулина на поступление в клетку глюкозы.
Б. Влияние на утилизацию глюкозы. Как показано ниже, инсулин оказывает влияние на внутриклеточную утилизацию глюкозы различными путями.
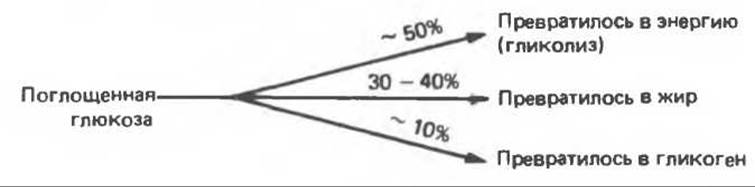
В норме примерно половина поглощенной глюкозы вступает на путь гликолиза и превращается в энергию, другая половина запасается в виде жиров или гликогена. В отсутствие инсулина ослабевает интенсивность гликолиза и замедляются анаболические процессы гликогенеза и липогенеза. Действительно, при инсулинодефицитном диабете всего лишь 5% поглощенной глюкозы превращается в жир.
Инсулин усиливает интенсивность гликолиза в печени, повышая активность и концентрацию ряда ключевых ферментов, таких, как глюкокиназа, фосфофруктокиназа и пируваткиназа. Более интенсивный гликолиз сопровождается более активной утилизацией глюкозы и, следовательно, косвенно способствует снижению выхода глюкозы в плазму. Инсулин, кроме того, подавляет активность глюкозо-6-фосфатазы — фермента, обнаруживаемого в печени, но не в мышцах. В результате глюкоза удерживается в печени, так как для глюкозо-6-фосфата плазматическая мембрана непроницаема.
В жировой ткани инсулин стимулирует липогенез путем 1) притока ацетил-СоА и NADPH, необходимых для синтеза жирных кислот, 2) поддержания нормального уровня фермента ацетил-СоА-карбоксилазы, катализирующего превращение ацетил-СоА в малонил-Соа, и 3) притока глицерола, участвующего в синтезе триацилглицеролов. При инсулиновой недостаточности все эти процессы ослабляются и в результате интенсивность липогенеза снижается. Другой причиной снижения липогенеза при инсулиновой недостаточности служит тот факт, что жирные кислоты, высвобождающиеся в больших количествах под действием некоторых гормонов, не встречающих противодействия со стороны инсулина, подавляют собственный синтез, ингибируя ацетил-СоА-карбоксилазу. Из всего сказанного следует, что суммарный эффект влияния инсулина на жир — анаболический.
Механизм влияния инсулина на утилизацию глюкозы включает в себя и другой анаболический процесс. В печени и в мышцах инсулин стимулирует превращение глюкозы в глюкозо-6-фосфат, который затем подвергается изомеризации в глюкозо-1-фосфат и в таком виде включается в гликоген под действием фермента гликогенсинтазы (ее активность также стимулируется инсулином). Это действие имеет двойственный и непрямой характер. Инсулин снижает внутриклеточный уровень сАМР, активируя фосфодиэстеразу. Поскольку сАМР-зависимое фосфорилирование инактивирует гликогенсинтазу, при низком уровне этого нуклеотида фермент находится в активной форме. Инсулин активирует и фосфатазу, катализирующую дефосфорилирование гликогенсинтазы, тем самым активируя этот фермент. И наконец, инсулин ингибирует фосфорилазу с помощью механизма, работающего с участием сАМР и фосфатазы, как описано выше. В результате высвобождение глюкозы из гликогена снижается. Таким образом, влияние инсулина на метаболизм гликогена также является анаболическим.
В. Влияние на образование глюкозы (глюконеогенез). Влияние инсулина на транспорт глюкозы, гликолиз и гликогенез проявляется за считанные секунды или минуты, поскольку первичные реакции этого влияния сводятся к активации или инактивации ферментов путем их фосфорилирования или дефосфорилирования. Более продолжительное влияние инсулина на содержание глюкозы в плазме крови связано с ингибированием глюконеогенеза. Образование глюкозы из предшественников неуглеводной природы осуществляется в результате ряда ферментативных реакций, многие из которых стимулируются глюкагоном (действие которого опосредовано сАМР), глюкокортикоидными гормонами и в меньшей степени а- и ß-адренергическими агентами — ангиотензином II и вазопрессином. Инсулин же подавляет эти ферментативные реакции. Роль ключевого фермента глюконеогенеза в печени принадлежит фосфоенолпируват-карбоксикиназе (ФЕПКК), катализирующей превращение оксалоацетата в фосфоенолпируват. Недавние исследования (см. ниже) показывают, что под действием инсулина количество этого фермента снижается в результате избирательного ингибирования транскрипции гена, кодирующего мРНК для фосфоенолпируват-карбоксикиназы.
Г. Влияние на метаболизм глюкозы. Результирующее действие всех перечисленных выше эффектов инсулина сводится к снижению содержания глюкозы в крови. Этому действию инсулина противостоят эффекты целого ряда гормонов, что, несомненно, отражает один из важнейших защитных механизмов организма, поскольку длительная гипогликемия способна вызвать несовместимые с жизнью изменения в мозге и, следовательно, ее нельзя допускать.
Д. Влияние на метаболизм липидов. Липогенное действие инсулина уже рассматривалось в разделе, посвященном его влиянию на утилизацию глюкозы. Кроме того, инсулин является мощным ингибитором липолиза в печени и жировой ткани, оказывая, таким образом, непрямое анаболическое действие. Частично это может быть следствием способности инсулина снижать содержание сАМР (уровень которого в тканях повышается под действием липолитических гормонов глюкагона и адреналина), а также способности инсулина ингибировать активность гормон-чувствительной липазы. В основе такого ингибирования лежит, по-видимому, активация фосфатазы, которая дефосфорилирует и тем самым инактивирует липазу или сАМР-зависимую протеинкиназу. В результате инсулин снижает содержание жирных кислот в крови. Это в свою очередь вносит вклад в действие инсулина на углеводный обмен, поскольку жирные кислоты подавляют гликолиз на нескольких этапах и стимулируют глюконеогенез. Данный пример показывает, что при обсуждении регуляции метаболизма нельзя учитывать действие лишь какого-либо одного гормона или метаболита. Регуляция — сложный процесс, в котором превращения по определенному метаболическому пути представляют собой результат сложных взаимодействий целого ряда гормонов и метаболитов.
У больных с инсулиновой недостаточностью активность липазы повышается, что приводит к усилению липолиза и увеличению концентрации жирных кислот в плазме и печени. Содержание глюкагона у таких больных также повышается, и это тоже усиливает выход свободных жирных кислот в кровь. (Глюкагон оказывает противодействие многим эффектам инсулина, и метаболический статус при диабете отражает соотношение уровней глюкагона и инсулина). Часть свободных жирных кислот метаболизируется до ацетил-СоА (обращение липогенеза) и затем в лимоннокислом цикле — до СO2 и Н2O. При инсулиновой недостаточности емкость этого процесса быстро оказывается превышенной и ацетил-СоА превращается в ацетоацетил-СоА и затем в ацетоуксусную и ß-гидроксимасляную кислоты. Под действием инсулина происходят обратные превращения.
Инсулин, по-видимому, влияет на образование или клиренс липопротеинов очень низкой плотности и липопротеинов низкой плотности, поскольку у больных с плохой компенсацией диабета содержание этих частиц, а следовательно, и содержание холестерола часто бывает повышенным. Именно этот метаболический дефект лежит, очевидно, в основе такого серьезного осложнения, как ускоренный атеросклероз, наблюдаемый у многих больных диабетом.
Влияние инсулина на метаболические процессы проиллюстрировано на рис. 51.14, где изображен ряд важнейших метаболических превращений в отсутствие инсулина.
Е. Влияние на метаболизм белков. Инсулин, как правило, оказывает анаболическое действие на белковый обмен, поскольку он стимулирует синтез белков и уменьшает их распад. Инсулин стимулирует поглощение мышцей нейтральных аминокислот типа А — эффект, не связанный с поглощением глюкозы или с последующим включением аминокислот в белки. Влияние инсулина на синтез белков в скелетной и сердечной мышцах проявляется, по-видимому, на уровне трансляции мРНК.
В последние годы было показано, что инсулин влияет на синтез специфических белков, вызывая изменения в соответствующих мРНК. Возможно, именно этим объясняется действие гормона на активность или количество отдельных белков. (Подробнее эта проблема обсуждается ниже.)
Ж. Влияние на размножение клеток. Инсулин стимулирует пролиферацию ряда клеток в культуре. Возможно, он участвует и в регуляции роста in vivo. При изучении регуляции роста чаще всего используются культуры фибробластов. В таких клетках инсулин усиливает способность фактора роста фибробластов (ФРФ), тромбоцитарного фактора роста (ТФР), фактора роста эпидермиса (ФРЭ), стимулирующих рост опухолей форболовых эфиров, простагландини F2a (ПГF20), вазопрессина и аналогов сАМР активировать размножение клеток, остановленных в фазе G1 в результате удаления из среды сыворотки.
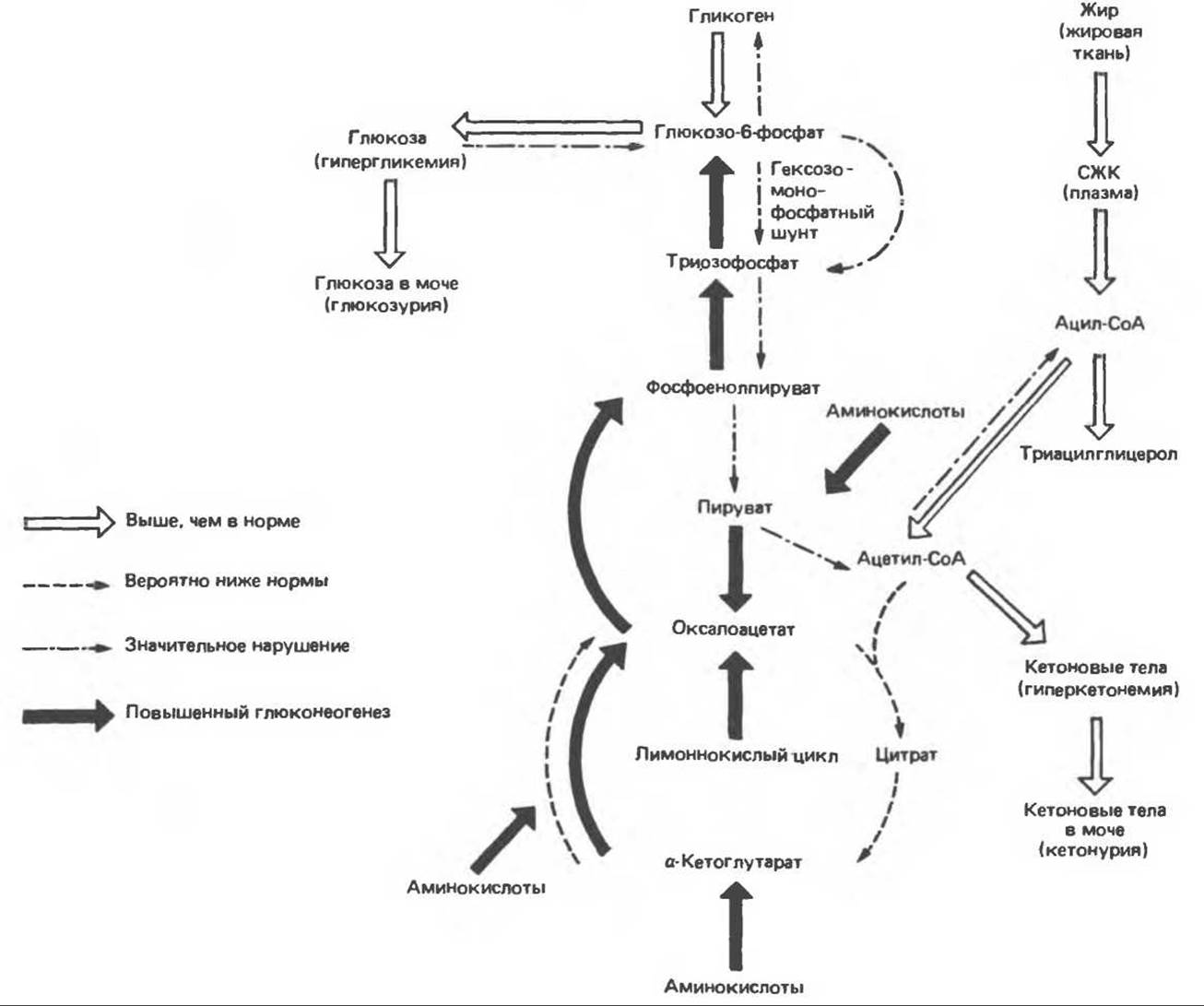
Рис. 51.14. Метаболические последствия инсулиновой недостаточности. СЖК — свободные жирные кислоты.
Преходящая потребность в различных факторах роста лежит в основе концепции о существовании двух классов таких факторов. Один из них, к которому относятся ТФР, ФРФ, ПГF2 и форболовые эфиры, вызывает, по-видимому, какие-то биохимические изменения в ранней G-фазе, которые, возникнув, устраняют дальнейшую потребность клетки в этих факторах и делают ее способной к репликации. Факторы роста второго класса (к ним относится инсулин) способствуют «продвижению» клетки к S-фазе и через нее и должны присутствовать постоянно. Данная модель описывает процессы, происходящие в фибробластах ЗТЗ, и ее универсальность не доказана. Не известно также, связан ли эффект инсулина с его взаимодействием с собственным рецептором или с рецептором инсулиноподобного фактора роста (ИФР) (тем более, что ИФР-1 тоже является фактором «продвижения»).
Инсулин поддерживает рост и репликацию многих клеток эпителиального происхождения, в том числе гепатоцитов, клеток гепатомы, клеток опухоли коры надпочечника и клеток карциномы молочной железы. Очень низкие концентрации инсулина стимулируют репликацию (по-видимому, через инсулиновый рецептор), причем нередко это происходит в отсутствие других пептидных факторов роста. Действительно, инсулин является необходимым компонентом всех известных сред для культивирования тканей, так что его значение для роста и репликации клеток несомненно.
Биохимический механизм влияния инсулина на репликацию клеток не выяснен; предполагают, что он основан на анаболическом действии гормона. Возможно, здесь играет роль влияние на поглощение глюкозы, фосфата, нейтральных аминокислот типа А и катионов. Гормон может стимулировать репликацию, используя свою способность активировать или инактивировать ферменты путем регуляции скорости и степени фосфорилирования белков или регулируя синтез ферментов.
Весьма интересная новая область исследований связана с изучением тирозинкиназной активности. Инсулиновый рецептор, как и рецепторы многих других факторов роста, включая ТФР и ФРЭ, обладает тирозинкиназной активностью. Важно отметить, что по крайней мере 10 онкогенных продуктов (многие из которых, вероятно, участвуют в стимулировании репликации злокачественных клеток) также представляют собой тирозинкиназы. Клетки млекопитающих содержат аналоги этих онкогенов (протоонкогены), продукты которых могли бы участвовать в репликации нормальных клеток. В пользу предположения о роли протоонкогенов свидетельствуют недавние работы, показавшие, что экспрессия, по крайней мере двух продуктов протоонкогенов — c-fos и c-mys, — после добавления сыровотки к культуре клеток с остановленным ростом усиливается. Показано также что, ТФР стимулирует образование специфических мРНК. Предстоит выяснить, аналогичен ли механизм действия инсулина.
Механизм действия инсулина
А. Рецептор инсулина. Действие инсулина начинается с его связывания со специфическим гликопротеиновым рецептором на поверхности клетки-мишени. Различные эффекты этого гормона (рис. 51.15) могут проявляться либо через несколько секунд или минут (транспорт, фосфорилирование белков, активация и ингибирование ферментов, синтез РНК), либо через несколько часов (синтез белка и ДНК и клеточный рост).
Инсулиновый рецептор подробно исследован с помощью биохимических методов и технологии рекомбинантных ДНК. Он представляет собой гетеродимер, состоящий из двух субъединиц (а и ß) в конфигурации a2-ß2, связанных между собой дисульфидными мостиками (рис. 51.15). Обе субъединицы содержат много гликозильных остатков. Удаление сиаловой кислоты и галактозы снижает как способность связывать инсулин, так и активность этого гормона. Каждая из гликопротеиновых субъединиц обладает особой структурой и определенной функцией. а-Субъединица (мол. масса 135000) целиком расположена вне клетки, и связывание инсулина, вероятно, осуществляется с помощью богатого цистином домена. ß-Субъединица (мол. масса 95000) — трансмембранный белок, выполняющий вторую важную функцию рецептора (гл. 44), т. е. преобразование сигнала. Цитоплазматическая часть ß-субъединицы обладает тирозинкиназной активностью и содержит участок аутофосфорилирования. Считается, что и то и другое важно для преобразования сигнала и действия инсулина (см. ниже). Поразительное сходство между тремя рецепторами, выполняющими различные функции, проиллюстрировано на рис. 51.16. Действительно, последовательности некоторых участков ß-субъединицы гомологичны таковым в рецепторе ФРЭ.
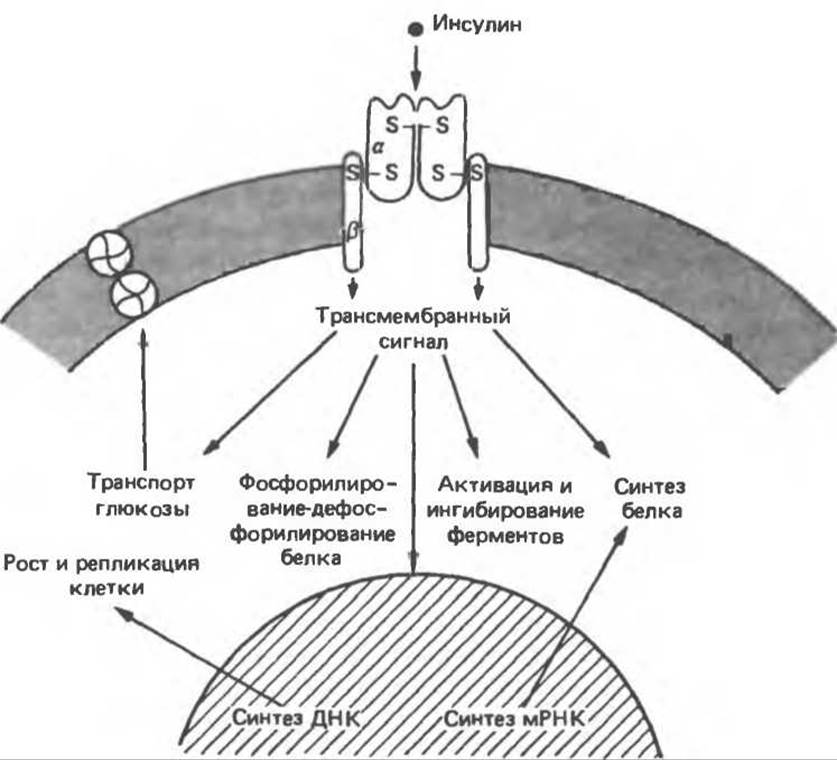
Рис. 51.15. Связь между рецептором инсулина и его действием. (Courtesy of С. R. Kahn.)
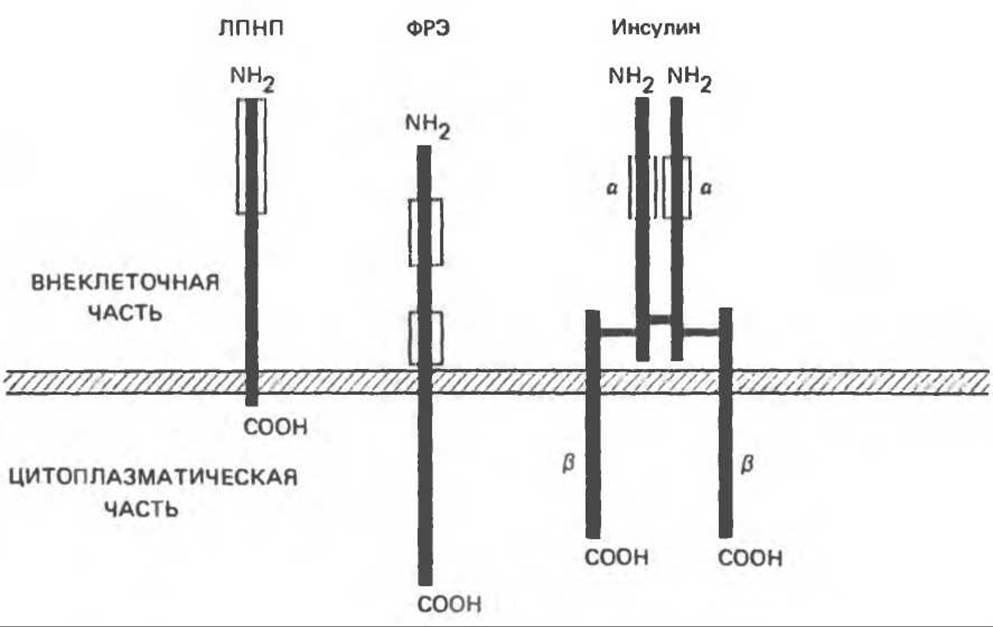
Рис. 51.16. Схема строения рецепторов липопротеинов низкой плотности (ЛПНП), фактора роста эпидермиса (ФРЭ) и инсулина. В каждом из этих рецепторов аминоконцы находятся в той части молекулы, которая выступает из клетки. Рамками обозначены участки, богатые цистеином, которые, как считают, участвуют в связывании лиганда. В каждом рецепторе (~25 аминокислот) имеется короткий домен, пересекающий плазматическую мембрану (заштрихованная полоса), и внутриклеточный домен варьирующей длины. Рецепторы ФРЭ и инсулина обладают тирозинкиназной активностью, локализованной в цитоплазматическом домене; кроме того, в этом домене находятся участки, в которых происходит аутофосфорилирование. Инсулиновый рецептор представляет собой гетеротетрамер, отдельные цепи (вертикальные полосы) которого связаны между собой дисульфидными мостиками.
Рецептор инсулина постоянно синтезируется и распадается; его период полужизни составляет 7— 12 ч. Рецептор синтезируется в виде одноцепочечного пептида в шероховатом эндоплазматическом ретикулуме и быстро гликозилируется в аппарате Гольджи. Предшественник человеческого рецептора инсулина состоит из 1382 аминокислот, его мол. масса составляет 190000, при расщеплении он образует зрелые а- и ß-субъединицы. У человека ген инсулинового рецептора локализован в хромосоме 19.
Рецепторы инсулина обнаружены на поверхности большинства клеток млекопитающих. Их концентрация достигает 20 000 на клетку, причем часто они выявляются и на таких клетках, которые не относят к типичным мишеням инсулина. Спектр метаболических эффектов инсулина хорошо известен. Однако инсулин участвует и в таких процессах, как рост и репликация клеток (см. выше), органогенез и дифференцировка у плода, а также в процессах заживления и регенерации тканей. Строение инсулинового рецептора, способность различных инсулинов связываться с рецепторами и вызывать биологические реакции практически идентичны в клетках всех типов и у всех видов. Так, свиной инсулин почти всегда в 10—20 раз эффективнее свиного проинсулина, который в свою очередь в 10—20 раз эффективнее инсулина морской свинки даже у самой морской свинки. Инсулиновый рецептор имеет, по-видимому, высоко консервативную структуру, еще более консервативную, чем структура самого инсулина.
При связывании инсулина с рецептором происходят следующие события; 1) изменяется конформация рецептора, 2) рецепторы связываются друг с другом, образуя микроагрегаты, пятна (patches) или нашлепки, 3) рецептор подвергается интернализации и 4) возникает какой-то сигнал. Значение конформационных изменений рецептора не известно, но интернализация, вероятно, служит средством регуляции количества и кругооборота рецепторов. В условиях высокого содержания инсулина в плазме, например при ожирении или акромегалии, число инсулиновых рецепторов снижается и чувствительность тканей-мишеней к инсулину уменьшается. Такая «снижающая» регуляция обусловлена потерей рецепторов в результате их интернализации, т. е. процесса проникновения инсулин-рецепторных комплексов в клетку путем эндоцитоза с помощью покрытых клатрином пузырьков (см. гл. 41). «Снижающая» регуляция объясняет отчасти инсулинорезистентность при ожирении и сахарном диабете II типа.
Б. Внутриклеточные медиаторы. Хотя механизм действия инсулина изучается более 60 лет, некоторые важнейшие вопросы, например природа внутриклеточного сигнала, остаются нерешенными, и инсулин в этом отношении не исключение. Внутриклеточные посредники не идентифицированы для очень многих гормонов (табл. 44.1). Множество различных молекул рассматривалось в качестве возможных внутриклеточных вторых посредников или медиаторов. К ним относятся сам инсулин, кальций, циклические нуклеотиды (cAMP, cGMP), Н2О2, пептиды мембранного происхождения, фосфолипиды мембраны, одновалентные катионы и тирозинкиназа (рецептор инсулина). Не одно из предположений не подтвердилось.
В центре внимания современных исследователей лежит тот факт, что инсулиновый рецептор сам является ферментом, чувствительным к инсулину, поскольку при связывании инсулина он подвергается аутофосфорилированию. Эта функция осуществляется ß-субъединицей, которая, действуя как протеинкиназа, переносит у-фосфат с АТР на остаток тирозина в ß-субъединице. Инсулин повышает Vmах этой ферментативной реакции, а двухвалентные катионы, особенно Мn2+, снижают Км для АТР.
Фосфорилирование тирозина нетипично для клеток млекопитающих (на долю фосфотирозина приходится всего 0,03% фосфоаминокислот, содержащихся в нормальных клетках), и вполне возможно, что наличие у рецепторов ФРЭ, ТФР, ИФР-1 тирозинкиназной активности неслучайно. Существует предположение, что тирозинкиназная активность — важный фактор в действии продуктов ряда вирусных онкогенов. Их связь с клеточными аналогами онкогенов, обладающими сходными свойствами при злокачественном и нормальном клеточном росте, рассматривалась выше. Изучение структуры этих компонентов выявило высокую степень гомологии между рецепторами и онкогенами, например между рецептором ФРЭ и erb-В, между рецептором ТФР и v-sis и между инсулиновым рецептором и v-ros.
Участие тирозинкиназы в преобразовании инсулин-рецепторного сигнала не доказано, но оно могло бы заключаться в фосфорилировании специфического белка, инициирующего действие инсулина, в запуске каскада фосфорилирование-дефосфорилирование, в изменении некоторых свойств клеточной мембраны или образовании какого-то связанного с мембраной продукта, например фосфолипида.
В. Фосфорилирование-дефосфорилирование белка.
Многие из метаболических эффектов инсулина, особенно те, которые возникают быстро, опосредованы его влиянием на реакции фосфорилирования-дефосфорилирования белка, что в свою очередь влияет на ферментативную активность данного белка. Перечень ферментов, активность которых регулируется таким путем, приведен в табл. 51.3. В некоторых случаях инсулин снижает внутриклеточное содержание сАМР (активируя сАМР-фосфодиэстеразу), что приводит к уменьшению активности cAMP-зависимой протеинкиназы. Такие эффекты характерны для гликогенсинтазы и фосфорилазы. В других случаях действие инсулина не зависит от сАМР и сводится к активации других протеинкиназ (например, в случае тирозинкиназы инсулинового рецептора), ингибированию третьих протеинкиназ (табл. 44.4) или (что значительно чаще) к стимуляции фосфатаз фосфопротеинов. Дефосфорилирование увеличивает активность ряда ключевых ферментов (табл. 51.3). Такие ковалентные модификации обеспечивают почти мгновенные изменения активностей ферментов.
Таблица 51.3. Ферменты, степень фосфорилирования и активность которых регулируются инсулином. (Modified and reproduced, with permission, from Denton R. M. et al: a partial view of the mechanism of insulin action. Diabetologia 1981, 21, 347.)
|
Фермент |
Изменения активности |
Возможный механизм |
|
Метаболизм с АМР Фосфодиэстераза (низкая Км) |
Повышается |
Фосфорилирование |
|
Протеинкиназа (сАМР-зависимая) |
Понижается |
Ассоциация R- и С-субъединиц |
|
Метаболизм гликогена Гликогенсинтаза |
Повышается |
Дефосфорилирование |
|
Киназа фосфорилазы |
Понижается |
» |
|
Гликолиз и глюконеогенез |
||
|
Пируватдегидрогeназа |
Повышается |
» |
|
Пируваткиназа |
» |
» |
|
6-Фосфофрукто-2-киназа |
» |
» |
|
Фруктозо-2,6-бисфосфатаза |
Понижается |
» |
|
Обмен липидов Ацетил-СоА — карбоксилаза |
Повышается |
Фосфорилирование |
|
ГМГ-СоА-редуктаза (гидроксиметилглутарил-СоА-редуктаза) |
» |
Дефосфорилирование |
|
Триацилглицерол-липаза |
Понижается |
» |
|
Другие процессы Тирозинкиназа (рецептор инсулина) |
? |
Фосфорилирование |
Г. Влияние на трансляцию мРНК. Известно, что инсулин влияет на количество и активность по крайней мере 50 белков в различных тканях, причем многие из этих эффектов сводятся к ковалентной модификации. Представление о роли инсулина в трансляции мРНК основывается главным образом на данных о рибосомном S6-белке — компоненте рибосомной субъединицы 40S. Такой механизм мог бы обеспечивать общее влияние инсулина на синтез белков в печени, скелетных и сердечных мышцах.
Д. Влияние на экспрессию генов. Все описанные эффекты инсулина реализуются на уровне плазматической мембраны или в цитоплазме. Однако инсулин способен влиять (по-видимому, через свой внутриклеточный медиатор) и на некоторые специфические ядерные процессы. Фермент фосфоенолпируват-карбоксикиназа (ФЕПКК) катализирует скорость-лимитирующую реакцию глюконеогенеза. Синтез ФЕПКК под действием инсулина снижается, а следовательно, уменьшается и интенсивность глюконеогенеза. Сравнительно недавно было показано, что при добавлении инсулина к культуре клеток гепатомы уже через несколько минут избирательно уменьшается скорость транскрипции гена ФЕПКК (рис. 51.17). В результате уменьшается количество как первичного транскрипта, так и зрелой мРНКФЕПКК, что в свою очередь приводит к снижению синтеза ФЕПКК. Этот эффект проявляется при физиологических концентрациях инсулина (10-12—10-9 моль/л), опосредуется инсулиновым рецептором и, по-видимому, обусловлен снижением скорости синтеза мРНКФЕПКК.
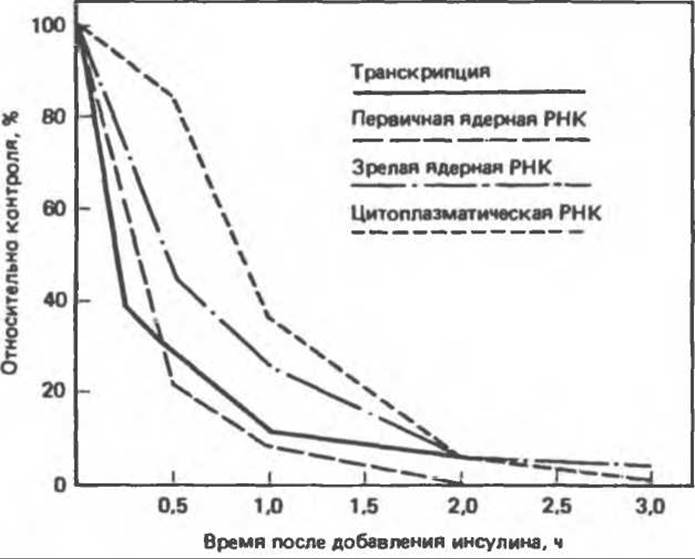
Рис. 51.17. Влияние гена инсулина на транскрипцию специфического гена. При внесении инсулина в культуру клеток гепатомы Н4ІІЕ скорость транскрипции гена ФЕПКК быстро снижается, что сопровождается уменьшением количества первичного транскрипта в зрелой мРНКФЕПКК. При уменьшении количества цитоплазматической мРНКФЕПКК снижается и скорость синтеза ФЕПКК-белка. (Reproduced, with permission, from Sasaki К. et al. Multihormonal regulation of phosphoenolpyruvate carboxykinase gene transcription, J. Biol. Chem., 1984, 259, 15242.)
Впервые влияние инсулина на транскрипцию генов было обнаружено при изучении механизма регуляции ФЕПКК, однако в настоящее время известны и другие примеры. Более того, представляется вероятным, что регуляция синтеза мРНК — главный эффект инсулина. Инсулин оказывает влияние на синтез многих специфических мРНК (табл. 51.4), в том числе на пока не идентифицированные мРНК в печени, жировой ткани и в мышцах (скелетных и сердечной). Доказано действие инсулина на транскрипцию генов овальбумина, альбумина и казеина.
Действие инсулина распространяется на ферменты, остающиеся в клетках, на секретируемые ферменты и белки, на белки, принимающие участие в процессах размножения, и на структурные белки (табл. 51.4). Эти эффекты регистрируются во многих органах и тканях и у многих видов. Регуляция транскрипции специфических мРНК под действием инсулина в настоящее время не вызывает сомнений. Этот путь модуляции ферментативной активности по важности не уступает механизму фосфорилирования- дефосфорилирования. Именно влиянием инсулина на транскрипцию генов, вероятно, объясняется его роль в эмбриогенезе, дифференцировке, а также росте и делении клеток.
Таблица 51.4. Белки, информационные РНК которых регулируются инсулином
|
Внутриклеточные ферменты Тирозин-аминотрансфераза1) Фосфоенолпируват-карбоксикиназа 1) Синтаза жирных кислот Пируваткиназа Глицерол-3-фосфат-дегидрогеназа1) Глицеральдегид-1-дегидрогеназа1) Глюкокиназа |
|
Секретируемые белки и ферменты Альбумин1) Амилаза а2-Глобулин Гормон роста1) |
|
Белки, участвующие в процессах размножения Овальбумин1) Казеин 1) |
|
Структурные белки 5-Кристаллин |
|
Другие белки В печени (р33 и т. п.) В жировой ткани В сердечной мышце В скелетных мышцах |
1) Инсулин регулирует скорость транскрипции соответствующих генов.
Патофизиология
При недостаточности инсулина или устойчивости к его действию развивается сахарный диабет. Примерно у 90% больных диабетом наблюдается инсулин-независимый сахарный диабет II типа (ИНЗСД). Для таких больных характерны ожирение, повышенное содержание в плазме инсулина и снижение количества инсулиновых рецепторов. У остальных 10% больных наблюдается диабет типа I, т.е. инсулинзависимый сахарный диабет I типа (ИЗСД). Рассмотренные выше метаболические нарушения более типичны именно для диабета типа I.
Ряд редких состояний иллюстрирует важные особенности действия инсулина. У некоторых людей образуются антитела к рецепторам инсулина. Эти антитела предотвращают связывание инсулина с рецептором, и в результате у таких лиц развивается синдром тяжелой инсулинорезистентности (см. табл. 43.2). При опухолях из В-клеток возникает гиперин-сулинемия и синдром, характеризующийся тяжелой гипогликемией. О важной роли инсулина (или, возможно, ИФР-1 или ИФР-2) для органогенеза свидетельствуют редкие случаи карликовости. Этот синдром характеризуется низким весом при рождении, малой мышечной массой, малым количеством подкожного жира, очень мелкими чертами лица, инсулинорезистентностью со значительным повышением содержания биологически активного инсулина в плазме и ранней смертью. У некоторых таких больных либо совсем отсутствовали рецепторы инсулина, либо они были дефектными.