Биохимия человека Том 2 - Марри Р. 1993
Биохимия внутри- и межклеточных коммуникаций
Мембраны: структура, сборка и функции
Передача информации в клетку
Специфические соединения, играющие роль биохимических сигналов, — нейромедиаторы, гормоны и иммуноглобулины — связываются с особыми рецепторами (интегральными белками), экспонированными с наружной стороны клеточной мембраны, и передают информацию через нее в цитоплазму. Например, ß-адренергический рецептор, который стереоспецифически связывает катехоламины, располагается на поверхности клеток-мишеней. Связывание с ним катехоламинов стимулирует активность аденилатциклазы, локализованной с внутренней стороны мембраны и катализирующей образование сАМР из АТР (гл. 44). Таким образом, информация, носителем которой во внеклеточной среде являлся специфический катехоламин, оказывается перенесенной внутрь, и ее последующую передачу осуществляет второй посредник, сАМР. Сопряженная с рецептором аденилатциклазная система, содержащая стимулирующие и ингибирующие компоненты, опосредует ответ на многие гормоны; более подробно это описано в гл. 44.
В клетках млекопитающих недавно был обнаружен другой тип передачи сигнала. В этой сигнальной системе роль второго посредника играет инозитолтрифосфат (рис. 42.21); его внутриклеточная концентрация регулируется внеклеточными сигналами, опосредованными трансмембранным рецептором. На поверхности большинства клеток млекопитающих располагаются специфические рецепторы для целой группы белков — факторов роста, таких, как инсулин, эпидермальный фактор роста и фактор роста, происходящий из тромбоцитов. При связывании соответствующей молекулы эффектора с рецептором на цитоплазматической стороне мембраны стимулируется киназная активность, присущая интегральному компоненту трансмембранной молекулы рецептора. Под действием этой активности происходит фосфорилирование фосфатид ил инозитола до фосфатидилинозитол-4-фосфата, а последнего до фосфатидилинозитол-4,5-бисфосфата. Интересно, что некоторые онкогены, экспрессия которых может приводить к малигнизации клеток, также индуцируют киназную активность, приводящую к образованию таких полифосфатидилинозитидов (гл. 57).
Другие рецепторы клеточной поверхности, например рецепторы для ацетилхолина, антидиуретического гормона и катехоламинов типа а1, при связывании с соответствующими лигандами могут способствовать активации фосфолипазы С. Последняя катализирует гидролиз фосфатидилинозитол-4,5-бисфосфата до инозитолтрифосфата и 1,2- диацилглицерола. Диацилглицерол способен активировать протеинкиназу С, активность которой зависит также от наличия в среде ионов Са2+. С другой стороны, инозитолтрифосфат приводит к эффективному высвобождению кальция из внутриклеточных депо, например саркоплазматического ретикулума и митохондрии. Таким образом, гидролиз инозитол-4,5-бисфосфата приводит к активации протеинкиназы С и содействует увеличению концентрации ионов кальция в цитоплазме. Это активирует Na+, K+-насос, ведет к суммарной утечке протонов из клетки и соответственно к увеличению внутриклеточного pH. В результате происходит пролиферация клетки и возникают другие специфические ответы. В этой сигнальной системе третьими посредниками, по-видимому, являются кальций и 1,2-диацилглицерол. Интересно, что на процессы, протекающие в этой сигнальной системе, влияют некоторые онкогены. Они опосредуют фосфатидилинозитолкиназную активность, что приводит к накоплению полифосфатидилинозитидов, которые в свою очередь служат предшественниками вторых и третьих посредников. По-видимому, будут обнаружены и другие сложные системы передачи информации в клетку. В гл. 44 обсуждается роль трансмембранных сигнальных систем в работе гормонов.
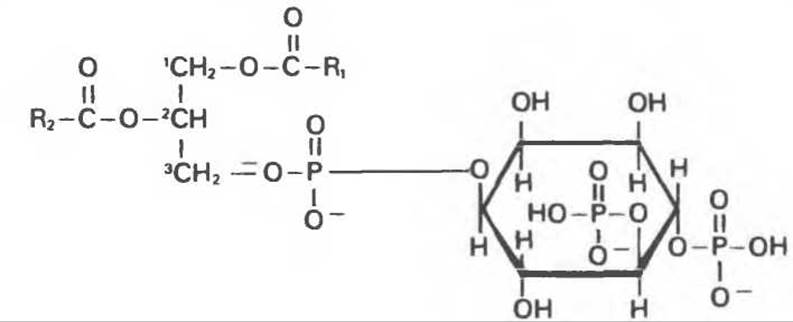
Рис. 42.21. Структура фосфатидилинозитол-4,5-бисфосфата.