Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Организация метаболизма: катаболические пути
Катаболизм сахаров
Пентозофосфатные пути
Важные метаболические пути, в которых участвуют пятиуглеродные пентозные сахара, называют либо пентозофосфатными путями, либо фосфоглюконатным путем, либо гексозомонофосфатным шунтом. Исторически первые данные о существовании таких путей были получены в экспериментах Варбурга по окислению глюкозо-6-фосфата в 6-фосфоглюконат. Напомним, что при изучении именно этой реакции был открыт NADP+ (гл. 2, разд. 3). Многие годы это окисление считали ферментативной реакцией, лежащей вне каких-либо определенных метаболических путей. Вместе с тем существовало предположение, что эта реакция является частью альтернативного пути распада глюкозы. Это предположение укрепилось после того, как было обнаружено, что процесс дыхания в тканях продолжается в присутствии высоких концентраций ионов фтора — известных ингибиторов енолазной реакции, — способных почти полностью блокировать процесс гликолиза. В некоторых тканях (в частности, в печени) этот альтернативный путь дыхания оказывается особенно активным. Теперь мы знаем, что пентозофосфатные пути многообразны и многоплановы. Они не только занимают существенное место в процессах катаболизма, но при функционировании в обратном направлении (восстановительный пентозофосфатный путь) являются ключевыми реакциями фотосинтеза, приводящими к образованию сахара
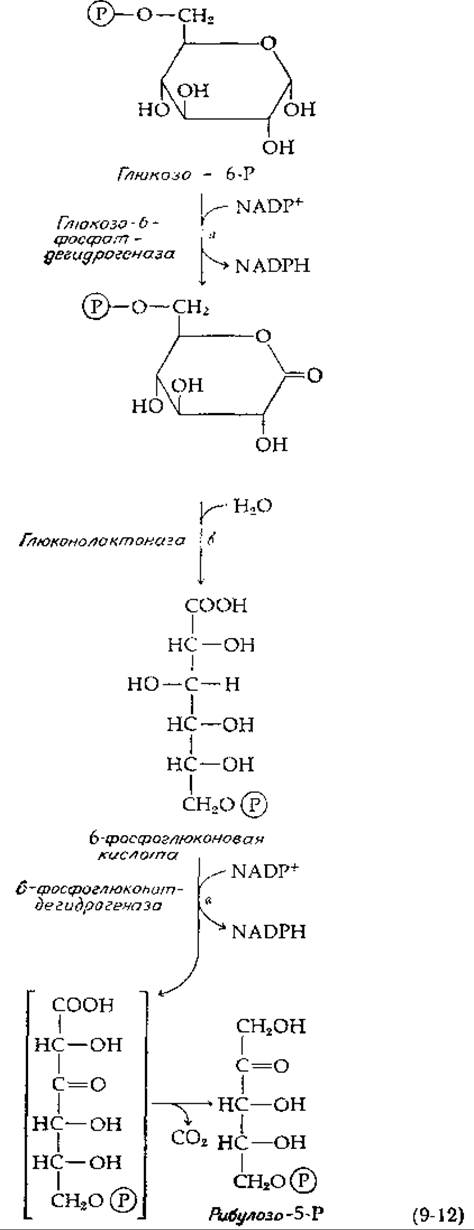
В ходе окислительных пентозных путей происходит отщепление от углеродной цепи сахара последовательно по одному атому углерода» который освобождается в форме СО2. Необходимые для этой цели ферменты составляют три различные системы; все они имеются в цитозоле живых клеток: I) дегидрогеназно-декарбоксилазная система, II) изомеризующая система и III) система структурной перестройки сахара. Дегидрогеназно-декарбоксилазная система расщепляет глюкозо-6-фосфат до СО2 и пентозофосфата [рибулозо-5-фосфат; уравнение (9-12)]. Для этого требуются три фермента, первым из которых является глюкозо-6-фосфат—дегидрогеназа [уравнение (9-12), стадия а; см. также уравнение (8-42)]. Непосредственный продукт реакции, лактон, может гидролизоваться самопроизвольно, однако глюконолактоназа [уравнение (9-12), стадия б] значительно ускоряет раскрытие кольца. Второе дегидрирование катализируется 6-фосфоглюконат-дегидрогеназой [уравнение (9-12), стадия в]; далее сразу же происходит катализируемое тем же ферментом ß-декарбоксилирование [по аналогии с уравнением (7-75)]. Значение ∆G0 для окисления глюкозо-6-фосфата в рибулозо-5-фосфат за счет NADP+ в соответствии с уравнением (9-12) составляет —30,8 кДж∙моль-1 — достаточнo большая отрицательная величина, чтобы в равновесном состоянии отношение [NADPH]/|NADP+] было выше 2000 при парциальном давлении СO2 в 0,05 атм.
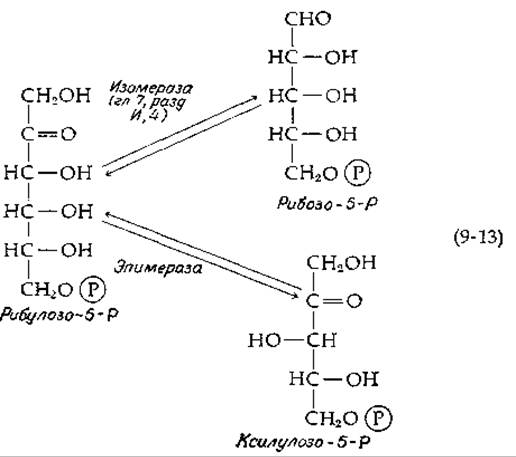
Изомеризующая система, состоящая из двух ферментов, обеспечивает взаимные превращения трех пентозофосфатов [уравнение (9-13); в приведенной формуле рибулозо-5-P опечатка, перешедшая из оригинала: одна СНОН-группа лишняя. — Ред.]. В результате образуется равновесная смесь этих трех пентозофосфатов. В дальнейших реакциях участвуют ксилулозо-5-фосфат и рибозо-5-фосфат.
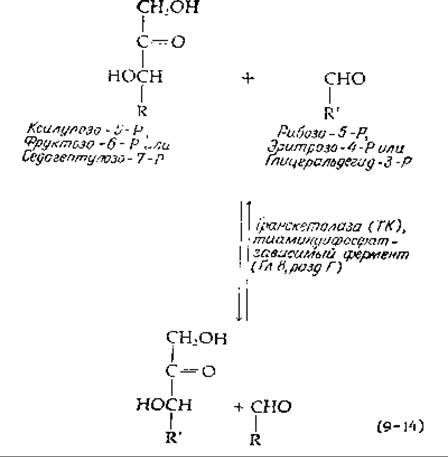
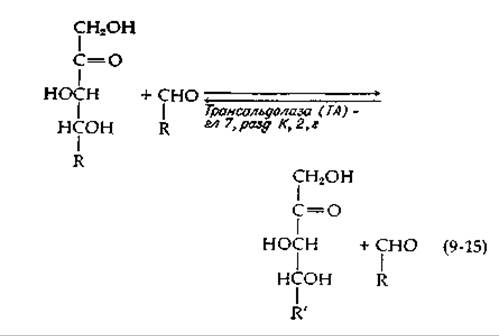
Система структурной перестройки сахара включает два фермента, транскетолазу и траисальдолазу. Оба фермента катализируют разрыв цепи и реакции переноса [уравнения (9-14) и (9-15)] у одной и той же группы субстратов. Эти ферменты осуществляют два основных типа разрыва связи С—С в двух положениях: по соседству с карбонильной группой (а) и у соседнего с карбонильной группой углеродного атома (ß). Как и в цикле трикарбоновых кислот, для пентозофосфатных путей оказываются необходимы разрывы цепи обоих типов.
а. Окислительный пентозофосфатный цикл
Соединив все три системы вместе, мы можем построить цикл окисления гексозофосфатов. В результате трех стадий декарбоксилирования (рис. 9-8,А) образуется трехуглеродный триозофосфат. Однако, поскольку дегидрогеназная система действует только на глюкозо-6-фосфат, в промежутках между тремя стадиями окисления должна функционировать система структурной перестройки. Обратите внимание, что C5-caxap (рибозо-5-фосфат), используемый в первой реакции, катализируемой транскетолазой, регенерируется в конце всей последовательности реакций. Таким образом, в этом цикле C5-caxap выступает в качестве регенерирующегося субстрата. Как указано штриховыми линиями, он может быть легко получен в любом требуемом количестве путем окисления глюкозо-6-фосфата. Прежде чем C5-caxap, образующийся на каждом этапе окисления, сможет быть вовлечен в реакции структурной перестройки, он должен быть изомеризован из рибулозо-5-фосфата в ксилулозо-5-фосфат; точно так же C5-caxap, образующийся на конечном этапе (рис. 9-8), может быть снова использован как регенерирующийся субстрат лишь после изомеризации в рибозо-5-фосфат. Система является весьма сложной. Отметим также, что на схеме, приведенной на рис. 9-8,А, в нескольких местах фигурируют одни и те же C5-субстраты. Таким образом, субстраты, образующиеся на разных стадиях цикла, перемешиваются, и вследствие этого процесс расщепления разных молекул гексозы складывается неодинаково.
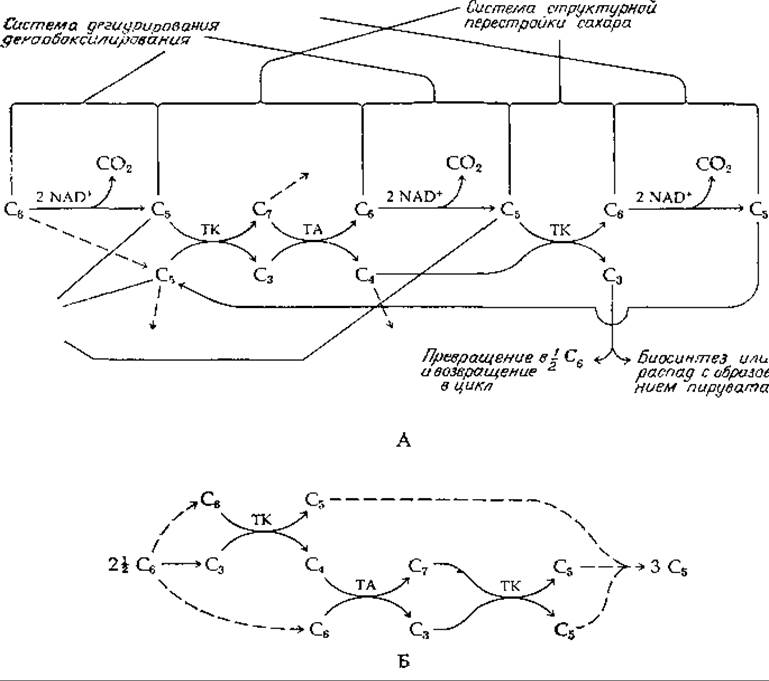
РИС. 9-8. Пентозофосфатные пути. А. Окисление гексозы (С6) до трех молекул СО2 и трехуглеродного фрагмента с возможностью вступления С3-, С4- и С7-продуктов в процессы биосинтеза (штриховые стрелки). Б. Неокислительные пентозные пути 21/2C6→3C5или 2С6→ЗС4, или 31/2С6→3С7.
Пентозофосфатный цикл часто рассматривают как процесс полного окисления гексоз в СО2. Чтобы осуществить такое окисление, С3-молекулы, рассматриваемые на рис. 9-8, А как продукты, должны быть превращены обратно в глюкозо-6-фосфат (под действием альдолазы, фосфатазы и гексофосфат-изомеразы), который снова вступает в цикл. Однако имеются и другие пути расщепления С3-продукта — фосфоглицеринового альдегида. Например, под действием ферментов гликолиза он может быть окислен до пирувата, а далее в цикле трикарбоновых кислот до СО2.
Как правило, в катаболических реакциях участвует NAD+, и поэтому не совсем обычно, когда в таких реакциях в качестве окислителя выступает NADP+. Тем не менее у млекопитающих ферменты пентозофосфатного цикла специфичны к NADP+. Существует предположение, что это связано с потребностью в NADPH для процессов биосинтеза (гл. 11, разд. В). Тогда становится понятным функционирование пентозофосфатного пути в тканях с наиболее активным биосинтезом (печень, молочная железа). Возможно, что в этих тканях С3-продукты цикла вовлекаются в процессы биосинтеза, как показано на рис. 9-8, А. Далее читателю должно быть уже понятно, что любой продукт от С4 до С7 может быть выведен из цикла в любых желаемых количествах без каких-либо нарушений в работе этого цикла. Например, мы знаем, что образующийся на промежуточной стадии С4-продукт эритрозо-4-фосфат используется бактериями и растениями (но не животными) для синтеза ароматических аминокислот. Подобным же образом рибозо-5-фосфат необходим для образования нуклеиновых кислот и некоторых аминокислот.
б. Неокислительные пентозофосфатные пути
Система структурной перестройки сахаров совместно с гликолитическими ферментами (превращающими глюкозо-6-фосфат в глицеральдегид-3-фосфат) может осуществлять превращение гексозофосфатов в пентозофосфаты (рис. 9-8,5) [34]. Полный процесс описывается уравнением
![]()
Читатель легко сможет показать, что те же ферменты могут катализировать превращение гексозофосфата в эритрозо-4-фосфат и в седогептулозо-7-фосфат:
![]()
Исследование метаболизма краснух липидобразующих дрожжей Rhodotorula gracilis (которые не содержат фосфофруктокииазы и поэтому неспособны к расщеплению сахаров по гликолитическому пути) свидетельствует о том, что 20% глюкозы окисляется по пентозофосфатному пути, тогда как 80% вовлекаются в реакции неокислительного пентозофосфатного пути1) [35, 36] [уравнение (9-16)]. Некоторые типы брожения также связаны с пентозофосфатными путями (разд. Е,6).
Оценка вкладов гликолитического, пентозофосфатного и других метаболических путей в энергетический баланс клетки сопряжена со многими трудностями, хотя попытки получить такие оценки продолжают предприниматься [35, 36].
1) Все же остается загадкой, откуда в условиях, когда блокирован гликолиз, берутся С3-молекулы, необходимые для функционирования иеокислительиого пути (рис. 9-8,5)? Вероятно, может происходить дальнейший распад пеитозофосфата на трехуглеродиые и двухуглеродиые единицы.