Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Ферменты: белковые катализаторы клеток
Специфичность действия ферментов
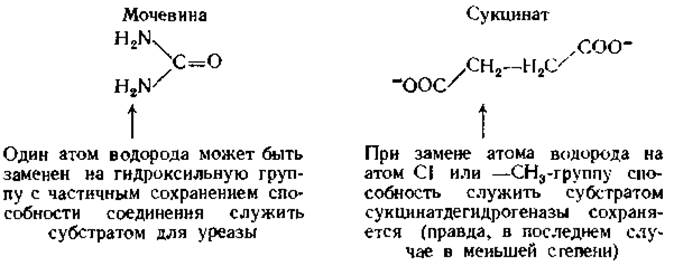
Одним из наиболее удивительных свойств ферментов является их высокая специфичность. В некоторых случаях специфичность по отношению к субстрату практически абсолютна. В течение многих лет полагали, что единственным субстратом для фермента уреазы является мочевина, а единственным субстратом для сукцинатдегидрогеназы — сукцинат. Даже после долгих поисков удалось найти лишь одно или два соединения с очень близкой структурой, на которые действовали указанное ферменты:
В других случаях ферменты проявляют специфичность к определенному классу соединений. Например, почечная оксидаза D-аминокислот катализирует окисление целого ряда D-аминокислот, но не действует на L-аминокислоты.
Не менее удивительна и специфичность ферментов к данному типу реакции. Довольно часто превращения субстрата могут идти по разным путям. Фермент катализирует только один путь, практически не влияя на другие реакции. Сопоставляя ферментативную реакцию с некатализируемой органической реакцией, поражаешься, что в то время как в ходе последней образуется большое количество побочных продуктов, ферментативная реакция протекает исключительно «чисто».
Дополнение 6-Б
Уреаза и следовые количества никеля
В дополнение к уже отмечавшейся практически абсолютной специфичности по отношению к своему субстрату (мочевине) уреаза, выделенная из канавалии мечевидной, примечательна еще тем, что она была первым ферментом, который удалось закристаллизовать. Это сделал в 1926 г. Дж. Самнер (J. Sumner). Хотя за это достижение Самнеру позднее была присуждена Нобелевская премия, его первые сообщения были встречены весьма скептически.
Уреаза катализирует гидролитическое расщепление мочевины с образованием двух молекул аммиака и молекулы углекислого газа и может использоваться для аналитического определения мочевины. Недавно было показано, что с каждой молекулой уреазы (мол. вес 105 000) связаны два атома никеля а-б. Наличие в молекуле уреазы нона металла не было ранее обнаружено, хотя на это указывало наличие в спектре поглощения очищенного фермента простирающегося в видимую область «хвоста» с плечом при 425 нм и небольшими максимумами при 725 и 1060 нм.
Согласно последним данным, никель необходим для животныхв. Так, на рационе, дефицитном по никелю, цыплята плохо росли, имели утолщенные ноги; у них наблюдались дерматиты. В тканях подобных животных обнаружены набухшие митохондрии и расширенное перинуклеарное пространство, что указывает на нарушение функции мембран. Токсичность никеля очень низка, и в организме животных существуют гомеостатические механизмы, регулирующие его концентрацию. Содержание этого элемента в тканях составляет от 1 до 5 мкг∙л-1. В сыворотке никель находится в составе низкомолекулярных комплексов, а также связан с сывороточным альбумином. Кроме того, известен специфический никельсодержащий белок класса макроглобулинов, названный никелоплазминомг. Никель присутствует в растениях, причем отдельные виды накапливают его в больших количествахд. Из-за повсеместной распространенности никеля составить рацион, в котором полностью отсутствовал бы этот элемент, довольно трудно.
Хотя никель может находиться в разных состояниях окисления, самым распространенным является Ni(II). Этот ион содержит восемь 3d-электронов, и поэтому координационное число равно четырем, причем лиганды располагаются в одной плоскости в вершинах квадрата. Однако ион Ni2+«амбивалентен»: он способен образовывать комплекс с шестью лигандами, обладающий октаэдрической структурой. Предполагается, что амбивалентность иона Ni2+имеет биохимическое значение. Какую роль играет ион Ni2+в функционировании уреазы, точно не установлено, однако не исключено, что он принимает участие в каталитическом процессе подобно иону Zn2+в карбоксипептидазе (рис. 7-3). Возможно также, что ион Ni2+образует координационное соединение с NH3 — продуктом расщепления субстрата. Высказывалось предположение, что ионы никеля или некоторых других переходных металлов содержат и ряд других ферментов, катализирующих гидролиз глутамина с образованием аммиака (гл. 14, разд. В, 2)6.
а Dixon N. Е., Gazzola С., Blakeley R. L., Zerner В. (1975). J. Amer. Chem. Soc., 97, 4131—4133.
б Dixon N. E., Gazzola C., Blakeley R. L., Zerner B. (1976). Science, 191, 1144—1150.
в Nielson F. H. (1974). In: Trace element metabolism in animals-2 (W. G. Hoekstra, J. W. Suttie, H. E. Ganther and W. Mertz, eds.), pp. 381—395. University Park Press, Baltimore, Maryland; см. также Schnegg A., Kichgessner M. (1976). Int. J. Vitamins Nutr. Res., 46, 96—99.
г Nomoto S., McNeely M. D., Sunderman F. W., Jr. (1971). Biochemistry, 10, 1647—1651.
д Severne В. C. (1974). Nature (London), 248, 807—808.