Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Биосинтез; как образуются новые молекулы
Стероидные соединения
Стероидные гормоны
В организме животных из холестерина образуются три важные группы гормонов: прогестины, половые гормоны и гормоны коры надпочечников (кортикостероиды). Основные пути биосинтеза этих гормонов показаны на рис. 12-17. Укорочение боковой цепи до двух углеродных атомов происходит путем гидроксилирования ее и последующего окислительного расщепления. В результате получается двухуглеродная боковая цепь, характерная для прегненолона — основного промежуточного продукта биосинтеза стероидов и кортикостероидов. Окисление 3-ОН-группы прегненолона в С = O сопровождается перемещением двойной связи; продуктом этой кетостероид-изомеразной реакции является a,β-ненасыщенный кетон — прогестерон [уравнение (7-56), реакция б].
а. Прогестины
Прогестерон — это основной гормон желтого тела, эндокринной железы, развивающейся в яичнике из фолликула после высвобождения яйцеклетки (овуляции). Прогестерон образуется также в надпочечниках, семенниках и плаценте. Он подвергается чрезвычайно быстрым превращениям в организме, в основном путем восстановления в соответствующий спирт, который затем образует конъюгированные соединения и выводится в виде глюкуронидов [уравнение (12-12)]. Восстановление двойной связи в кольце А прогестерона приводит к полной потере биологической активности, что указывает на важную роль а,β-ненасыщенного кетона в проявлении гормонального действия. Прогестерон функционирует как гормон, необходимый для поддержания беременности, и, кроме того, наряду с эстрогенами он участвует в регуляции менструального цикла.
б. Кортикостероиды
В коре надпочечников (т. е. в наружной зоне надпочечников) прогестерон превращается в две группы гормонов, важнейшими представителями которых являются кортизол и альдостерон. Секреция кортизола надпочечниками регулируется гормоном гипофиза кортикотропином (АКТГ, АСТН); секреция кортизола достигает у взрослого человека 15—30 мг в день. В крови кортизол присутствует в основном в связанной с белком форме; белок плазмы, транспортирующий кортизол, называется транскортином. Как упоминалось в гл. 11, разд. Е, 2, кортизол — это глюкокортикоид, стимулирующий глюконеогенез и накопление глюкогена в печени. В мышцах и других тканях кортизол ингибирует синтез белка, а в жировой ткани усиливает расщепление жиров с освобождением жирных кислот.
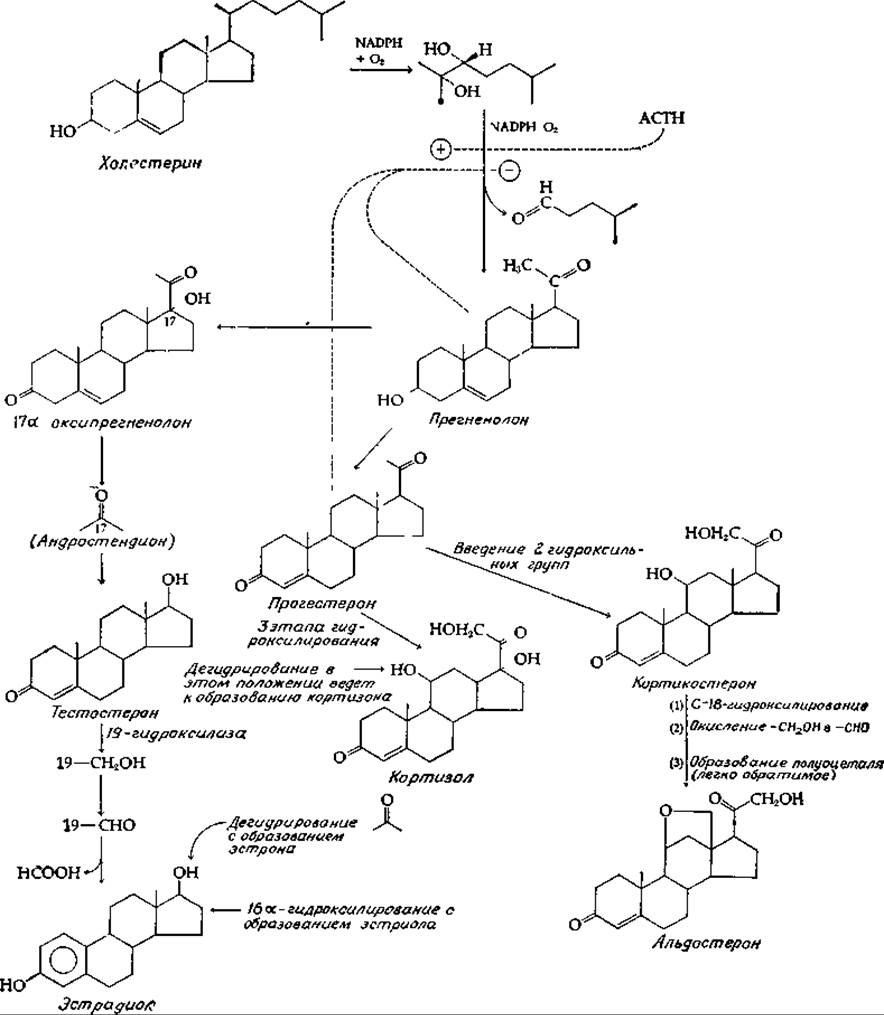
РИС. 12-17. Пути биосинтеза некоторых стероидных гормонов.
Широко известно антивоспалительное действие кортизола, а также близкого ему по структуре кортизона. В основе этого эффекта лежит способность указанных гормонов стабилизировать мембраны лизосом и усиливать распад лимфоцитов.
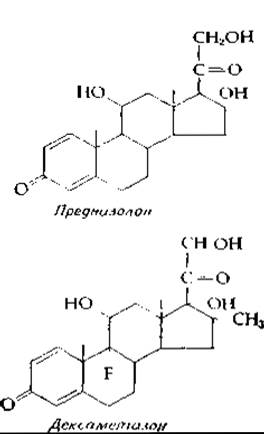
Кортизон и его синтетические аналоги, такие, как преднизолон или дексаметазон, принадлежат к числу современных «чудодейственных средств». Их применяют при острых приступах артрита, при тяжелых воспалениях глаз и других органов. Однако продолжительное использование этих препаратов может вызвать тяжелые побочные явления, в частности атрофию мышц и резорбцию костей. Последнее возникает в результате специфического торможения абсорбции кальция в желудочно-кишечном тракте под влиянием указанных средств. Таким образом, в этом отношении глюкокортикоиды являются антагонистами витамина D (дополнение 12-Г).
Образование альдостерона, относящегося к группе минералокортикоидов, регулируется системой ренин — ангиотензин. Эта гормональная система активируется при нарушении ионного баланса, выявляемого рецепторами ионов натрия в почках. Альдостерон усиливает обратное всасывание ионов натрия в почечных канальцах и таким образом регулирует водный и солевой обмен. Секреция альдостерона у взрослого человека при нормальном содержании натрия в пище составляет около 0,1—0,2 мг/день. Глюкокортикоиды также обладают небольшой минералокортикоидной активностью; так, обнаружилось, что больных с недостаточностью функции надпочечников (болезнь Аддисона) можно поддерживать на одних глюкокортикоидах (без минералокортикоидов) при адекватном потреблении соли с пищей.
Дополнение 12-Г
Витамин D
Недостаточность витамина D приводит к рахиту — заболеванию животных и человека, которое характеризуется размягчением и деформацией костей и их недостаточной кальцификациейa. Возникновение рахита как следствия дефицита какого-то компонента пищи признавалось уже свыше ста лет назад, а использование жира тресковой печени для предотвращения этого заболевания началось примерно в 1870 г. В 1890 г. была замечена связь рахита с недостаточным пребыванием на солнечном свету. Однако только в 1924 г., когда Стинбок и Хесс (Steenbock, Hess) показали, что облучение светом некоторых продуктов питания повышает их защитную противорахитную активность, витамин D (кальциферол) стали рассматривать как второй жирорастворимый витамин. Витамин D — это семейство соединений, образующихся при облучении светом ∆5,7-ненасыщенных стеринов, таких, как эргостерин и 7-дегидрохолестеринб. Из первого образуется эргокальциферол (витамин D2), из второго — холекальциферол (витамин D3).
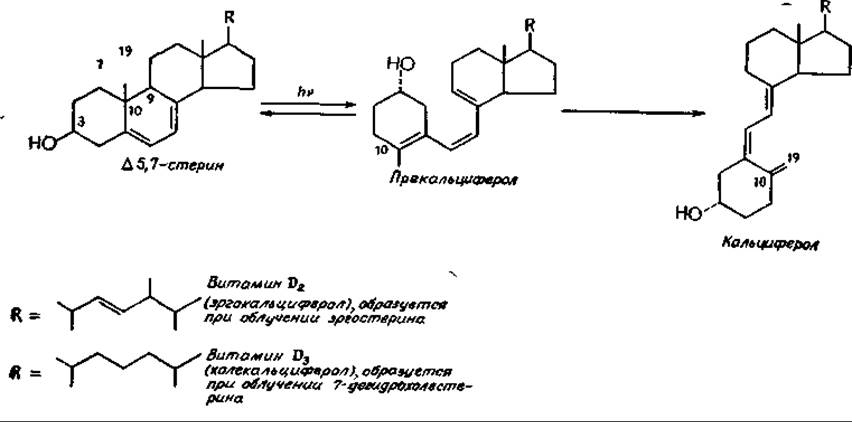
При низкой температуре удается выделить промежуточный продукт — прекальциферол, поскольку при этом в ходе фотохимической реакции устанавливается стационарная концентрация прекальциферола. При более высоких температурах последний превращается в кальциферол. Несколько медленнее протекают побочные фотохимические реакции, в ходе которых образуются другие соединения, в том числе токсические. Вследствие этого облучение светом эргостерина для использования в качестве пищевых добавок должно производиться с осторожностью.
В нормальных условиях человек получает достаточно солнечных лучей для образования нужных количеств холекальциферола из 7-дегидрохолестерина в коже, так что добавки витамина D в пищу не требуется. Это в особенности относится ко взрослым. Детям обычно рекомендуется давать с пищей ~20 мкг (400 М. Ед.) эргокальциферола в день. Большие количества витамина D нежелательны, а десятикратное увеличение указанной дозы оказывает значительный токсический эффект.
Основная функция витамина D состоит в регуляции обмена кальция. В последнее время было обнаружено, что собственно регуляторами обмена выступают полярные гидроксилированные метаболиты витамина D. Гидроксилирование протекает по трем положениям, и самым полярным из известных в настоящее время метаболитов является 1,24,25-триоксихолекальциферолв,г.
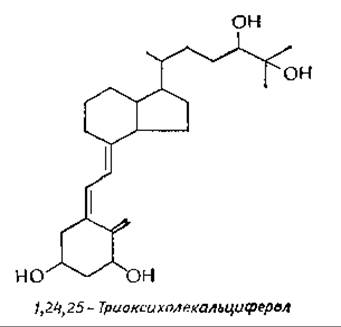
Эти метаболиты правильнее рассматривать как стероидные гормоны, а сам витамин D — как гормон, образующийся в коже. Изученные реакции гидроксилирования витамина D суммированы в приводимой ниже схеме. Заслуживает упоминания, что первое гидроксилирование с образованием 25-оксихолекальциферола протекает в печени, тогда как два последующих — в почках. Поскольку собственно регуляторами метаболизма ионов кальция служат ди- и триоксипроизводные витамина D, у больных с поражением почек часто наблюдается значительная деминерализация костей (почечная остеодистрофия). В последнее время проводятся исследования, направленные на получение синтетических полигидроксилированных производных витамина D для введения больным с поражением почекд.
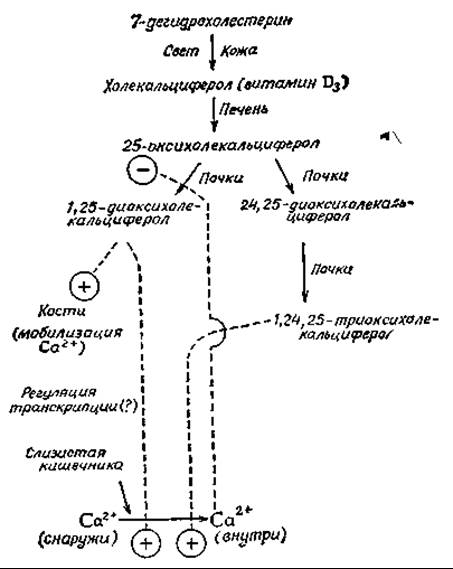
Как 1,25-диоксихолекальциферол, так и триоксипроизводное увеличивают всасывание ионов кальция в слизистой кишечника. По-видимому, эти соединения, подобно другим стероидным гормонам, осуществляют свое действие на уровне регуляции транскрипциие. Так, например, было показано, что в ответ на введение витамина D увеличивается содержание кальцийсвязывающих белков, которые, по всей вероятности, участвуют в процессе всасывания в кишечнике кальцияв,ж,з.
Соотношение концентраций образующихся в организме 1.25-диоксикальциферола и 24,25-производного зависит от ряда условий. В частности, наличие ионов кальция оказывает ингибирующее действие на 1-гидроксилазу, как это показано на приведенной схеме11. При этом эффект ионов кальция может быть как прямым, так и опосредованным действием паратгормонак. Если 1,24,25-триоксивитамин D оказывает специфическое воздействие на слизистую кишечника, то 1,25-диоксипроизводное влияет также на клетки кости, где вызывает мобилизацию ионов кальция. Этот эффект частично обусловлен стимуляцией Са-активируемой АТРазы наружных мембран клеток костии. 1,25-диоксивитамин D увеличивает также реабсорбцию неорганического фосфата в почкахл. Продукты метаболизма витамина D оказывают влияние также на мышцы и другие ткани.
а Сходное заболевание у крупного рогатого скота — «молочная лихорадка».
б Dyke S. F., The Chemistry of the Vitamins, pp. 271—317, Wiley (Interscience), New York, 1965.
в DeLuca H. F., Schnoes H. K., Annu. Rev. Biochem., 45, 631—666 (1976).
г Holick M. F., Kleiner-Bossaller A., Schnoes H. K., Kasten P. M., Boyle I. T., DeLuca H. F., JBC, 248, 6691—6696 (1973).
д Holick M. F., DeLuca H. F., Kasten P. M., Korycka M. B., Science, 180, 964—966 (1973).
е Procsal D. A., Okamura W. H., Norman A. W., JBC, 250, 8382—8388 (1975).
ж DeLuca H. F., Fed. Proc. Fed. Am. Soc. Exp. Biol., 33, 2211—2219 (1974).
з Emtage J. S., Lawson D. E. M., Kodicek E., Nature (London), 246, 100—101 (1973).
и Galante L., Colston К. W., Evans I. M. A., Byfield P. G. H., Matthews E. W., MacIntyre L, Nature (London), 244, 438—440 (1973).
к Russel R. G. G., Monod A., Bonjour J.-P., Fleisch H., Nature (London), New Biol., 240, 126—127 (1972).
л Tanaka Y., DeLuca H. F., PNAS, 71, 1040—1044 (1972).
в. Андрогены
Основной андроген, или мужской половой гормон, — это тестостерон, образующийся из прегненолона путем отщепления боковой цепи при С-17. Образование тестостерона составляет у мужчин 6—10 мг/сут, у женщин также синтезируется тестостерон, но в небольших количествах (~0,4 мг). Любопытно, что тестостерон служит предшественником женских половых гормонов. В крови тестостерон циркулирует в комплексе с ß-глобулином и оказывает воздействие на ряд органов-мишеней, в том числе и на органы воспроизведения. Наиболее заметный эффект тестостерона — это стимуляция роста волос на лице. Вместе с тем тестостерон вызывает преждевременную гибель волосяных фолликул на голове у генетически предрасположенных индивидуумов. В результате у лысых мужчин обычно растет прекрасная борода, причем волосяные фолликулы бороды при пересадке на темя остаются устойчивыми к действию андрогенов. Чем обусловлены различия этих регуляторных эффектов, совершенно неизвестно. Облысение можно вылечить, используя соответствующие антагонисты андрогенов, но при этом происходит выпадение бороды и пропадает половая активность. Одна из задач химиков-фармакологов — создание соединения, которое было бы способно блокировать действие андрогенов только на волосяные фолликулы головы.
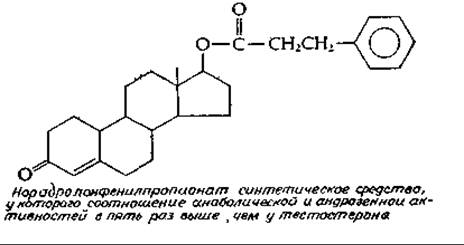
Андрогены обладают также общей «анаболической» активностью, проявляющейся в стимуляции синтеза белка, особенно в мышечной ткани. Они способствуют росту костей; бурный рост мальчиков и девочек в период полового созревания обусловлен, по всей видимости, именно андрогенами. Более высокий рост мужчин по сравнению с женщинами также связан с большим образованием андрогенов Попытки создания «анаболических гормонов», лишенных андрогенной активности, привели к появлению множества синтетических стероидов. Некоторые из этих препаратов, по крайней мере частично, обладали желательными свойствами, в настоящее время анаболические гормоны широко применяются спортсменами, хотя целесообразность этого мероприятия представляется весьма спорной.
г. Эстрогены
17ß-эстрадиол является основным эстрогенным, или женским, половым гормоном. Он образуется из тестостерона путем окислительного отщепления атома С-19 и последующей ароматизации кольца А[100]. Наличие этого ароматического кольца характерно для всех эстрогенных гормонов. Эстрогены синтезируются главным образом в яичниках, а во время беременности и в плаценте. В определенном количестве синтез идет и в семенниках. Так, семенники жеребца содержат больше эстрогенов, чем любая другая эндокринная железа. К тканям-мишеням для эстрогенов относятся прежде всего молочные железы и матка, а также и многие другие ткани организма. Эстрогены тормозят рост костей. Более гладкая кожа женщин и большее содержание в женском организме жировой клетчатки также обусловлены влиянием эстрогенов.
Эстрадиол и прогестерон — два гормона, участвующие в регуляции менструального цикла. В начале цикла образование обоих гормонов невелико. Далее происходит активация синтеза эстрогенов под влиянием фолликулостимулирующего гормона (ФСБ), вырабатываемого передней долей гипофиза и усиливающего рост в яичниках граафовых фоликулов, в которых синтезируются эстрогены. Примерно в середине цикла под влиянием лютеинизирующего гормона (ЛГ), также вырабатываемого гипофизом, происходит овуляция (освобождение яйцеклетки), после чего начинает продуцироваться прогестерон. Этот гормон необходим для поддержания беременности. Если имплантация бластодита не имела места, то образование прогестерона уменьшается и цикл заканчивается менструацией.
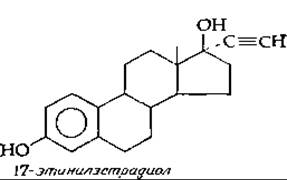
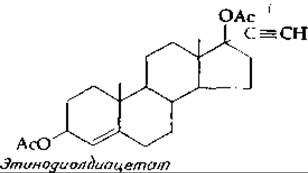
Введение эстрогенов и прогестинов угнетает секрецию ФСГ и ЛГ гипофизом (принцип отрицательной обратной связи) и тем самым прекращает процесс овуляции. Этот эффект лежит в основе действия контрацептивных (противозачаточных) препаратов. Прием препаратов может осуществляться по такой схеме: в течение первых 10—15 дней ежедневно принимают небольшие количества синтетического эстрогена 17-этинилэстрадиола, а затем в течение 10—15 дней тот же эстроген в комбинации с прогестином, например этинодиолдиацетатом. Согласно другой схеме, в течение всего времени следует принимать только прогестин.
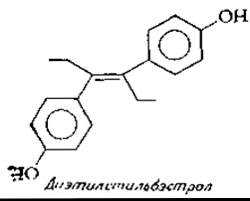
Одним из синтетических соединений, обладающим эстрогенной активностью, является диэтилстильбэстрол. Некоторое время его широко использовали при откорме скота для увеличения привеса. Затем было обнаружено, что при скармливании крысам больших количеств диэтилстильбэстрола проявляется канцерогенное действие препарата, в связи с чем его применение было прекращено. Представляется также сомни тельным использование диэтилстильбэстрола в форме таблеток для употребления «утром следующего дня» в качестве препарата, препятствующего имплантации оплодотворенной яйцеклетки.
Следует упомянуть о существовании растительных флавоноидов, обладающих эстрогенной активностью.