Биохимия - Химические реакции в живой клетке Том 2 - Д. Мецлер 1980
Ферменты: белковые катализаторы клеток
Механизмы ферментативного катализа
Эффекты сближения и ориентации
Несмотря на большие успехи, достигнутые энзимологией, и тот факт, что благодаря рентгеноструктурным исследованиям мы знаем структуру некоторых ферментов, в действии этих удивительных катализаторов остается еще много загадочного. Ни кислотно-основный, ни ковалентный катализ, по-видимому, не могут объяснить огромного увеличения скорости реакции, которое наблюдается, когда в работу вступают ферменты. Какие же другие факторы обусловливают высокую скорость катализируемых ферментами реакций?
Одно из самых первых предположений состояло в том, что ферменты просто сближают реагенты и удерживают их в таком положении достаточно долго, чтобы реакционноспособные группы могли столкнуться и в конце концов прореагировать. Интуитивно ясно, что фактор сближения должен играть очень важную роль, но ранние попытки количественно оценить этот эффект приводили к заключению, что роль его невелика. Позднее Пейдж и Дженкс показали, что проведенные оценки неверны и что исключительно за счет уменьшения энтропии двух реагентов при их сближении и удерживании на поверхности молекулы фермента можно ожидать увеличения скорости в 103 и более раз [60, 61]. Поскольку энтропия при связывании субстрата уменьшается, то становится ясным, что энтальпия этого процесса должна быть высока, а в таком случае связывание субстратов ферментом уже само по себе во многом определяет движущую силу катализа. Подобная идея была впервые высказана Вестхеймером [62], который считал, что ферменты за счет способности связывать субстрат создают как бы энтропийную ловушку.
Потеря энтропии поступательного и вращательного движения, которая, по оценкам Пейджа и Дженкса, составляет от —160 до —210 кДж∙моль-1∙град-1, перекрывает «неблагоприятную» энтропию активации, характерную для бимолекулярных реакций.
В последние годы больше внимания уделялось вопросу о том, насколько точной должна быть ориентация субстратов в случае быстрых реакций [63, 63а], и был исследован ряд модельных реакций, например спонтанное образование внутреннего эфира (лактона), протекающее с отщеплением воды:
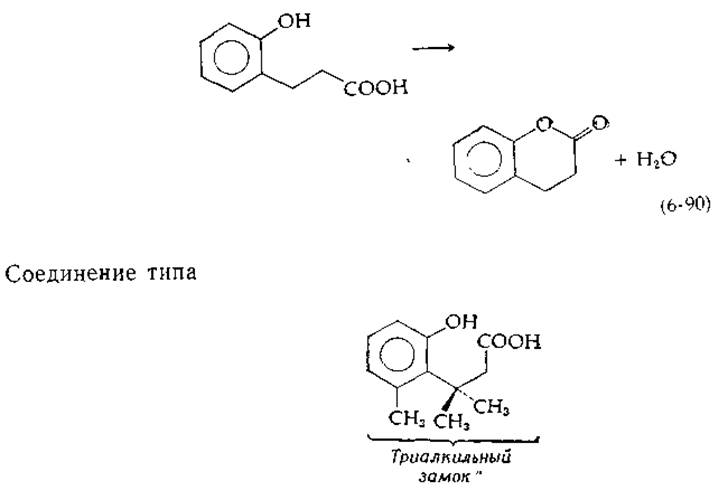
реагирует гораздо быстрее из-за существенных конформационных ограничений [64, 65]: скорость реакции в 10" раз выше скорости реакции (6-90), вероятно, вследствие того, что создаваемые ограничения приводят к существенному повышению частоты столкновения —СООН- и —ОН-групп. Три метильные группы сближаются и образуют триалкильный «замок». Эти результаты позволяют полагать, что ориентационные эффекты могут играть большую роль в ферментативном катализе.
Согласно другой концепции, объясняющей высокую скорость ферментативных реакций, ферменты способны индуцировать напряжение, или «искажение» в молекуле субстрата, приводящее к ослаблению специфических связей (см. гл. 7, разд. В, 4, а, посвященный лизоциму). Напряжение может либо сопровождаться конформационным изменением в самой белковой молекуле, либо возникать в результате подобного конформационного изменения. Еще один факт, который необходимо принять во внимание, состоит в том, что некоторые реакции протекают быстрее в среде с низкой диэлектрической постоянной, чем в воде. Возможно, полярные группы субстрата дегидратируются при связывании с активным центром фермента, и это приводит к повышению их реакционной способности.
У многих ферментативных реакций образование переходного состояния сопровождается существенным изменением объема (∆V≠). Возможно, эти изменения обусловлены в основном изменением степени гидратации групп, расположенных на поверхности молекулы фермента, и играют важную роль в ферментативном катализе [65а].