БИОХИМИЯ - Л. Страйер - 1984
ТОМ 1
ЧАСТЬ I. КОНФОРМАЦИЯ И ДИНАМИКА
ГЛАВА 10. ВВЕДЕНИЕ В ПРОБЛЕМУ БИОЛОГИЧЕСКИХ МЕМБРАН
Обратимся теперь к биологическим мембранам, которые представляют собой высокоорганизованные структуры, построенные главным образом из белков и липидов. Мембраны играют жизненно важную роль. Они отделяют клетки от окружающей среды, тем самым обусловливая их индивидуальность. Однако мембраны-это отнюдь не глухая преграда, а барьер с высокоизбирательной проницаемостью, в котором имеются специфические молекулярные насосы и каналы. Эти транспортные системы регулируют молекулярный и ионный состав внутриклеточной среды. В клетках эукариот имеются еще и внутренние мембраны, отграничивающие органеллы, например, митохондрии, хлоропласты, лизосомы. Функциональная специализация в процессе эволюции была тесно связана с формированием таких обособленных внутриклеточных участков, называемых обычно компартментами, или отсеками.
Мембраны регулируют также обмен информацией между клетками и средой. В частности, они несут специфические рецепторы, воспринимающие внешние стимулы. Движение бактерий к источнику пищи, ответ клетки-мишени на гормон (например, инсулин), восприятие света-все это примеры процессов, где в качестве первичного акта происходит распознавание сигнала специфическим рецептором на мембране. В свою очередь и некоторые мембраны сами способны генерировать сигнал-химический или электрический. Все это свидетельствует о том, что мембраны играют центральную роль в системе биологической коммуникации.
Два самых важных процесса превращения энергии в биологических системах протекают в мембранных системах, содержащих высокоупорядоченные наборы ферментов и других белков. Так, фотосинтез, т. е. процесс превращения света в энергию химической связи, происходит во внутренних мембранах хлоропластов (рис. 10.2), тогда как окислительное фосфорилирование, в ходе которого за счет окисления органических субстратов образуется аденозинтрифосфат (АТР), протекает во внутренних мембранах митохондрий. В последующих главах мы подробно рассмотрим эти, а также некоторые другие мембранные процессы. Настоящая глава посвящена некоторым наиболее важным свойствам, общим для большинства биологических мембран.
Рис. 10.2. Фотосинтезирующие комплексы тилакоидных мембран хлоропластов превращают свет в энергию химических связей

10.1. Общие свойства биологических мембран
Мембраны различаются как по функции, так и по структуре. Однако всем им присущи следующие основные свойства.
1. Мембраны представляют собой плоскую структуру толщиной в несколько молекул, образующую сплошную перегородку между отдельными отсеками (компартментами). Толщина мембран составляет обычно 60 100 А.
2. Мембраны состоят главным образом из липидов и белков. Весовое соотношение белков и липидов для большинства биологических мембран лежит в пределах от 1 :4 до 4 ; 1. В мембранах имеются также углеводные компоненты, связанные с липидами и белками.
3. Липиды мембран представлены относительно небольшими молекулами, несущими гидрофильные и гидрофобные группы. В водной среде эти липиды спонтанно образуют замкнутые бимолекулярные слои. Такие липидные двойные слои (бислои) служат барьером для полярных соединений.
4. Отдельные функции мембран опосредуются специфическими белками. Белки выполняют роль насосов, каналов, рецепторов, ферментов и преобразователей энергии. Белки мембран встроены (интеркалированы) в липидный бислой, что создает пригодную для проявления их активности среду.
5. Мембраны-нековалентные надмолекулярные структуры; составляющие мембрану белки и липиды удерживаются вместе благодаря возникновению множества нековалентных взаимодействий, кооперативных по своему характеру.
6. Мембраны асимметричны: их наружная и внутренняя поверхности отличаются друг от друга.
7. Мембраны - жидкие структуры. Если молекулы липидов, так же, как и белков, не зафиксированы в определенном месте силами специфического взаимодействия, то они легко диффундируют в плоскости мембраны. Мембраны можно рассматривать как двумерные растворы определенным образом ориентированных белков и липидов.
Рис. 10.3. Пространственные модели. А-пальмитат (С16, насыщенный); Б олеат (С18, ненасыщенный). Двойная связь цис-конфигурации обусловливает изгиб углеводородной цепи
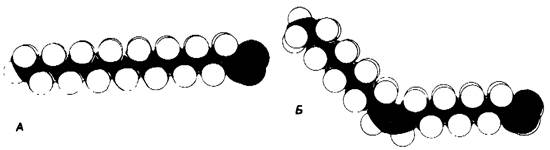
Рис. 10.4. Абсолютная конфигурация глицерол-3-фосфата в мембранных липидах. А-Н и ОН, присоединенные к С-2, находятся впереди плоскости рисунка, тогда как С-1 и С-3-позади нее. Б - изображение той же структуры по Фишеру; в этом случае горизонтальные линии связей соответствуют связям, выступающим из плоскости рисунка вперед, а вертикальные линии-связям, расположенным позади плоскости рисунка