БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
ЧАСТЬ V. МОЛЕКУЛЯРНАЯ ФИЗИОЛОГИЯ
ГЛАВА 36. МЕМБРАННЫЙ ТРАНСПОРТ
36.18. Через щелевые соединения ионы и небольшие молекулы перетекают из клетки в клетку
Большие водные каналы, через которые происходит пассивный транспорт, свойственны многим биологическим мембранам прокариот и эукариот. Так, в наружной мембране грамотрицательных бактерий имеются каналы, образованные порином — трансмембранным белком массой 37 кДа (разд. 32.14). По этим каналам диаметром 10 А легко проникают в периплазматическое пространство полярные молекулы массой, не превышающей примерно 600 Да. Далее эти молекулы транспортируются в цитозоль посредством симпорта, транслокации групп или иных пермеаз. Подобным же образом в наружных мембранах митохондрий и хлоропластов имеются большие водные каналы, образованные из аналогичных порину молекул.
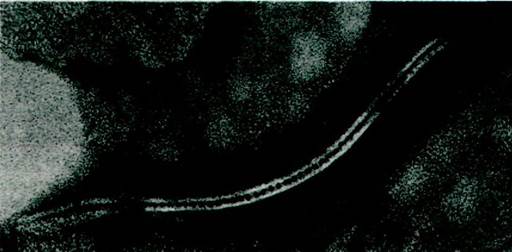
Наиболее хорошо изученным типом водного канала у эукариот является щелевое соединение (щелевой контакт), называемое также межклеточным каналом, поскольку он служит протоком между внутренним содержимым многих смежных клеток. Щелевые соединения, открытые Жан-Полем Ревелем и Морисом Карновским (Jean-Paul, Revel, Morris Kamovsky), располагаются скопом (кластерами) в определенных участках плазматических мембран прилежащих друг к другу клеток. На электронных микрофотографиях листков щелевых соединений (рис. 36.28) можно видеть, что каждое соединение образовано шестью субъединицами, окружающими пору диаметром 15-20 А. На тангенциальном срезе (рис. 36.29) видно, что эти гексамеры пронизывают промежуток (щель) между соприкасающимися клетками (отсюда и название - щелевое соединение). Для определения внутреннего размера щелевых соединений производили микроинъекцию ряда флуоресцирующих веществ в одни клетки и затем наблюдали, как распространяется флуоресценция по соседним клеткам. По данным Вернера Лёвенштейна (Werner Loewenstein), все полярные вещества массой до 1 кДа легко проходят по межклеточным каналам. Другими словами, по щелевым соединениям из одной клетки в другую могут поступать неорганические ионы и большинство метаболитов (сахара, аминокислоты, нуклеотиды). Что касается белков, нуклеиновых кислот и полисахаридов, то из-за своих больших размеров они не проходят по этим каналам.
Рис. 36.28. Электронная микрофотография изолированных листков межклеточных щелевых соединений. Цилиндрические коннексоны образуют гексагональную решетку с ребром ячейки 85 А. Диаметр интенсивно окрашенной полости в центре - около 20 А

Рис. 36.29. Электронная микрофотография тангенциального среза щелевых соединений между прилежащими клеточными мембранами
Щелевые соединения играют важную роль в межклеточной коммуникации. В ряде возбудимых тканей, например, в сердечной мышце, клетки объединены в единую систему быстрым потоком ионов через эти соединения; таким путем достигается быстрый и синхронный ответ на стимуляцию. Через щелевые контакты происходит также питание клеток, удаленных от кровеносных сосудов, например, в костях или хрусталике глаза. Представляется вероятным, что рассматриваемые каналы коммуникации имеют важное значение в регуляции процессов развития и дифференцировки.
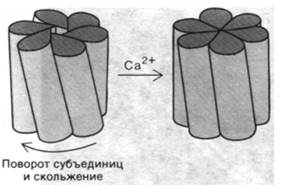
Проницаемость щелевых контактов регулируется ионами кальция. Повышение внутриклеточного содержания Са2+ приводит к тому, что щелевой контакт в той или иной мере закрывается. Межклеточные каналы полностью открыты при концентрации Са2+ ниже 10-7 М и полностью закрываются при концентрации Са2+, превышающей 5 • 10-5 М. Увеличение содержания Са2+ в указанном диапазоне приводит к сужению просвета межклеточных каналов, причем в первую очередь снижается проницаемость для более крупных молекул. Структурная основа таких изменений просвета каналов была выявлена при анализе реконструированных трехмерных изображений полей щелевых контактов, взятых в двух различающихся по четвертичной структуре состояниях. Эти структурные исследования показали, что щелевой контакт состоит из двух смыкающихся цилиндрических единиц, названных коннексонами. Каждый коннексон образован шестью субъединицами, имеет длину 75 А и насквозь пронизывает плазматическую мембрану. Сегмент коннексока длиной 20 А выступает во внеклеточное пространство и соединяется с коннексоном прилежащей клетки. Длинная ось каждой из субъединиц коннексона имеет наклон по отношению к поперечной оси мембраны. Путем скольжения субъединиц относительно друг друга происходит уменьшение их наклона к поперечной оси мембраны, и канал при этом закрывается (рис. 36.31). Самые большие конформационные изменения имеют место в середине канала, где субъединицы двух стыкующихся коннексонов скользят относительно друг друга на расстояние примерно 11 А, что соответствует повороту на 28°. Анализ высокой степени разрешения позволит в дальнейшем выявить, каким образом Са2+ индуцирует скольжение и поворот.
Рис. 36.30. Схематическое изображение щелевого контакта
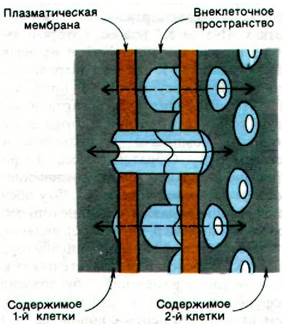
Рис. 36.31. Модель регуляции ионами Са2+ степени закрывания щелевого соединения