БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 25. ИНФОРМАЦИЯ РНК И ТРАНСКРИПЦИЯ
25.15. Матричная ДНК содержит стоп-сигналы для транскрипции
Терминирование транскрипции регулируется так же тонко, как и инициирование. В матричной ДНК имеются стоп-сигналы, которые были расшифрованы путем сравнения различных последовательностей оснований в этих участках. Все они имеют одну общую особенность: вслед за GC-богатой областью перед участком терминации располагается АТ-богатая последовательность. Отличительная особенность терминирующих последовательностей — симметрия второго порядка этой GC-богатой области (рис. 25.17). Следовательно, РНК-транскрипт этой области самокомплементарен, а это значит, что он способен к спариванию оснований, приводящему к образованию структуры шпильки (рис. 25.18). Кроме того, новообразованные цепи РНК кончаются несколькими остатками U, которые кодируются серией оснований А в АТ-богатой области ДНК-матрицы. Одна или несколько подобных структурных особенностей заставляют РНК-полимеразу задержаться, сделать паузу, когда она сталкивается с таким сигналом. В некоторых участках терминации новообразованные цепи РНК высвобождаются без участия дополнительных белков. В других местах для терминации цепи необходимо участие белка р (ро).
Рис. 25.17. Последовательность ДНК, соответствующая 3'-концу trp-мРНК E. coli. Последовательности оснований, показанные желтым цветом, обладают симметрией второго порядка относительно оси, обозначенной зеленым цветом.
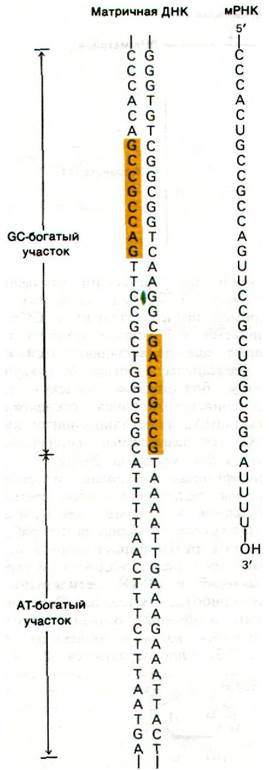
Рис. 25.18. Последовательность оснований З'-конца мРНК, транскрибированной с триптофанового оперона E. coli. Возможно образование стабильной структуры шпильки
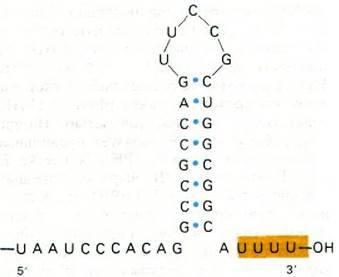
25.16. Белок р участвует в терминировании транскрипции
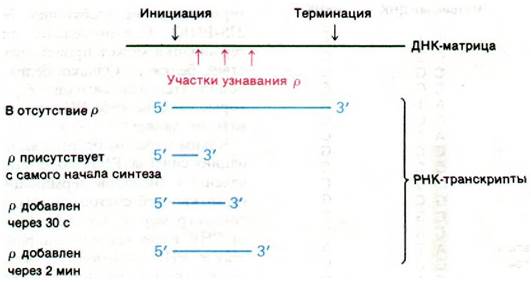
Тот факт, что терминирование транскрипции в некоторых участках происходит с участием белка р, подтверждается следующими данными: молекулы РНК, синтезированные in vitro в присутствии белка р, короче молекул, полученных в его отсутствие. Например, РНК, синтезированная на ДНК фага fd в присутствии белка р, имеет коэффициент седиментации 10S, тогда как РНК, синтезированная в отсутствие белка p, 23S. Дополнительные сведения о действии белка р были получены при добавлении этого фактора терминации в инкубационную смесь через различные промежутки времени после начала синтеза РНК. Если белок p добавляли через несколько секунд, через 2 и 10 мин после инициации, получали РНК с коэффициентами седиментации 13, 17 и 23S соответственно. Этот результат свидетельствует о том, что матрица содержит по крайней мере три участка терминации, чувствительных к белку p (они дают 10S, 13S- и 17S-РНК), и один участок терминации, не требующий белка p (дает 23S-PHK). Следовательно, специфическая терминация может происходить и в отсутствие белка р. Однако белок р выявляет дополнительные сигналы терминации, которые сама по себе РНК-полимераза узнавать не может.
Рис. 25.19. Влияние фактора р на размер транскрибируемых РНК
Каким образом белок р вызывает терминацию синтеза РНК по достижении определенных сигналов терминации? Одна из возможностей состоит в том, что этот белок-тетрамер с массой 200 кДа - садится на РНК и движется по направлению к молекуле РНК-полимеразы, останавливая ее на участке терминации. Согласно этой модели, белок р вытесняет РНК-полимеразу с 3'-конца РНК, что приводит к высвобождению РНК-транскрипта. Интересно отметить, что при терминировании транскрипции белок р гидролизует АТР.
Как можно было предвидеть, терминирование транскрипции некоторых генов контролируется. В ходе дальнейшего изложения мы увидим, что бактериофаг ![]() синтезирует белки-антитерминаторы, обеспечивающие возможность транскрипции и экспрессии некоторых генов (разд. 28.11), У Е. coli действует система регуляции с использованием специализированных терминирующих сигналов, называемых аттенюаторами, для обеспечения пищевых потребностей клетки (разд. 28.9).
синтезирует белки-антитерминаторы, обеспечивающие возможность транскрипции и экспрессии некоторых генов (разд. 28.11), У Е. coli действует система регуляции с использованием специализированных терминирующих сигналов, называемых аттенюаторами, для обеспечения пищевых потребностей клетки (разд. 28.9).
25.17. Многие молекулы РНК после транскрипции расщепляются и химически модифицируются
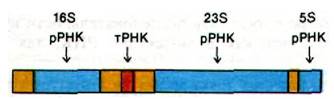
Образование функционально активных молекул РНК (процессинг) продолжается после завершения транскрипции. У прокариот молекулы транспортных и рибосомных РНК образуются путем расщепления и химической модификации определенных новосинтезированных цепей РНК. Например, у E. coli три вида молекул рибосомных РНК и одна молекула транспортной РНК вырезаются из первичного РНК-тран- скрипта, который, кроме того, содержит спейсерные (разграничивающие) участки (рис. 25.20). Другие транскрипты содержат по несколько различных видов тРНК или несколько копий одной и той же тРНК. Нуклеазы, которые расщепляют и укорачивают эти предшественники рРНК и тPHK, действуют с высокой точностью. Например, рибонуклеаза Р образует правильные 5'-концы всех молекул тРНК в клетке Е. coli. Рибонуклеаза III вырезает предшественники 5S-, 16S- и 23S-pPHK из первичного транскрипта, расщепляя определенные связи в двухспиральных шпилечных областях. Молекулы мРНК у прокариот, наоборот, практически не претерпевают модификации. Более того, многие из них транслируются еще до того, как заканчивается их транскрипция. В то же время некоторые вирусные мРНК (например, мРНК фага Т7) расщепляются рибонуклеазой III раньше, чем начинается трансляция.
Второй тип процессинга - присоединение нуклеотидов к концам некоторых РНК. Например, последовательность ССА присоединяется к 3'-концам молекул тРНК. которые еще не обладают этой концевой последовательностью. У эукариот к 3'-концу большинства молекул мРНК присоединяется длинная последовательность poly(A), а метилированный нуклеотид G (так называемый «колпачок», или «кеп») - к 5'-концу (разд. 29.22).
Рис. 25.20. При расщеплении этого первичного транскрипта образуются молекулы 5S, 168 и 23S рРНК и одна молекула тРНК. Спейсерные участки закрашены желтым цветом
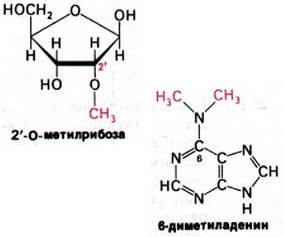
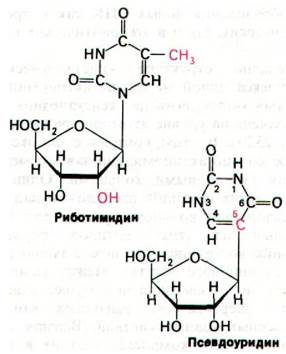
Модификации оснований и рибозных остатков представляют собой третий тип процессинга. У эукариот одна 2'-гидрок- сильная группа примерно на сто рибозных остатков в рРНК ферментативно метилируется за счет S-аденозилметионина. У бактерий в рРНК метилируются не остатки рибозы, а основания. Особенно интересны необычные основания, которые встречаются во всех молекулах тРНК (разд. 27.3). Они образуются путем ферментативной модификации обычных рибонуклеотидов, входящих в состав предшественника тРНК. Например, псевдоуридилат и риботимидилат образуются путем модификации уридиловых остатков после транскрипции.
У эукариот все транскрипты подвергаются интенсивному процессингу. Расщепление и модификация предшественников рРНК и тРНК напоминают соответствующие процессы у прокариот. Удивительное отличие состоит в том, что мРНК эукариот образуется путем расщепления большого транскрипта и последующего сращивания (сплайсинга) получающихся при этом фрагментов. Это явление мы рассмотрим в одной из следующих глав (разд. 29.21). У эукариот транскрипция и трансляция происходят в различных компартментах клетки. По всей вероятности, процессинг РНК играет ключевую роль в регуляции транспорта мРНК, рРНК и тРНК из ядра в цитозоль.