БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 26. ГЕНЕТИЧЕСКИЙ КОД Т ЗАВИСИМОСТЬ МЕЖДУ ГЕНАМИ И БЕЛКАМИ
26.10. Последовательность оснований гена и последовательность аминокислот соответствующего полипептида коллинеарны
Обратимся теперь к взаимоотношениям между генами и белками. Как показала работа Сеймура Бензера (Seymour Benzer) по генетическому картированию высокого разрешения, гены - неразветвленные структуры. Этот важный результат согласовывался с установленным фактом, что ДНК представляет собой линейную последовательность пар оснований. Полипептидные цепи также имеют неразветвленную структуру. Поэтому в начале 60-х годов возник следующий вопрос: существует ли линейное соответствие между геном и его полипептидным продуктом?
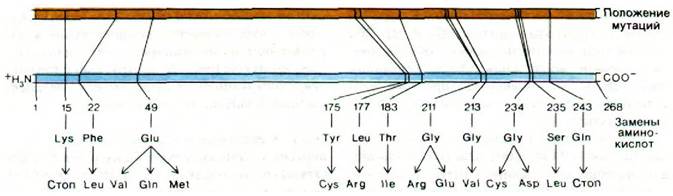
Подход Чарлза Янофски (Charles Yanofsky) к этой проблеме состоял в использовании мутантов E. coli, у которых образуется измененная молекула фермента. Было выделено много мутантов по a-цепи триптофан-синтазы и определена локализация мутаций на генетической карте a-цепи с помощью рекомбинационных экспериментов с трансдуцирующим фагом. Некоторые из этих мутаций были локализованы близко друг к другу на генетической карте, тогда как другие были сильно удалены в пределах одного гена. Следующей важной задачей было определить положение аминокислотной замены для каждого из этих десяти мутантов. Прежде всего была определена последовательность 168 аминокислот a-цепи дикого типа. Затем с помощью метода «отпечатков пальцев» были установлены место и природа аминокислотной замены в каждом случае. Порядок расположения мутаций на генетической карте был таким же, как порядок соответствующих замен в последовательности аминокислот полипептидного продукта (рис. 26.5). Другими словами, ген, кодирующий α-цепь, и его полипептидный продукт коллинеарны.
Рис. 26.5. Коллинеарность гена и аминокислотной последовательности a-цепи триптофан-синтазы. Положение мутаций в ДНК (желтая линия) было определено методами генетического картирования. Аминокислотные замены в последовательности аминокислот (синяя линия) расположены в том же порядке, что и соответствующие мутации
26.11. Некоторые последовательности вирусных ДНК кодируют более одного белка
Открытие того факта, что ДНК фага ɸХ174 кодирует больше белков, чем позволяет имеющееся в ней количество нуклеотидов, было совершенно загадочным. Как этот вирус может кодировать более 2000 аминокислотных остатков, если он содержит всего лишь 5375 нуклеотидов? Ответ был получен из полной последовательности оснований (разд. 24.29), когда обнаружилось, что в ДНК фага фХ174 некоторые гены перекрываются. Определенные участки соответствующих транскриптов транслируются в различных рамках считывания, что приводит к образованию белков с различными последовательностями аминокислот (рис. 26.6). Например, одни и те же 300 нуклеотидов кодируют белок гена Е и большую часть белка гена D. Еще более поразительный пример представляет родственный бактериофаг G4, в ДНК которого некоторые короткие участки кодируют три различных белка. Эти вирусы используют перекрывающиеся гены, чтобы ввести больше информации в маленькие молекулы ДНК. Однако за эту генетическую экономию приходится платить: на последовательности аминокислот, кодируемые перекрывающимися генами, накладываются строгие ограничения. Поэтому перекрывающиеся гены, по-видимому, широко используются лишь в тех случаях, когда количество ДНК строго лимитировано, как в случае вирусов с белковой оболочкой строго определенных размеров.
Рис. 26.6. Перекрывающиеся гены
в ДНК фага ɸХ174. Рядом с последовательностью оснований показаны две последовательности аминокислот, детерминируемые различными рамками считывания

26.12. Гены эукариот представляют собой мозаику из транслируемых и нетранслируемых последовательностей ДНК
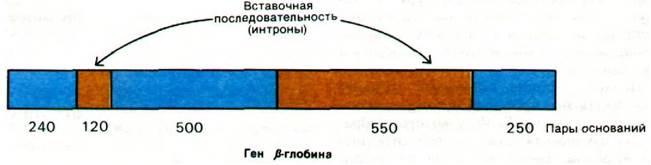
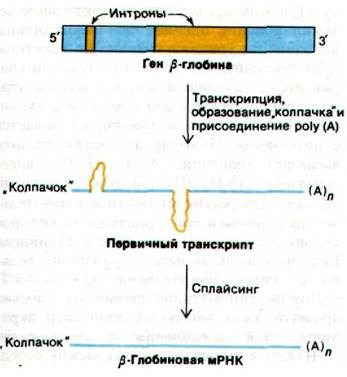
У бактерий полипептидные цепи кодируются непрерывной последовательностью триплетных кодонов. В течение многих лет считалось, что гены высших организмов также непрерывны. Эта точка зрения была неожиданно опровергнута в 1977 г., когда в нескольких лабораториях было открыто, что некоторые гены имеют прерывистое строение. Например, ген β-цепи гемоглобина прерывается в области, кодирующей аминокислотную последовательность, длинной некодирующей вставочной последовательностью из 550 пар оснований и короткой последовательностью из 120 пар оснований. Таким образом, ген β-глобина разделен на три кодирующие последовательности:
Эта удивительная структура была открыта с помощью электронно-микроскопических исследований гибридов между β-глобиновой мРНК и фаргментом ДНК мыши, содержащим ген β-глобина (рис. 26.7). Двухцепочечная ДНК частично денатурируется, что позволяет мРНК гибридизоваться с комплементарной цепью ДНК. Затем одноцепочечный участок ДНК образует петлю и на электронных микрофотографиях выглядит как тонкая линия в отличие от двухцепочечной ДНК или ДНК-РНК-гибридных участков, которые выглядят значительно толще. Если бы ген β-глобина был непрерывен, была бы видна одна петля. Однако на электронных микрофотографиях таких гибридов (рис. 26.8) отчетливо видны три петли. Это показывает, что ген прерывается по крайней мере одним участком ДНК, которого нет в соответствующей мРНК. Дополнительные данные о вставочных последовательностях были получены при картировании рестрикционных фрагментов гена β-глобина и продукта обратной транскрипции мРНК. Большие различия между этими картами показали, что геномная ДНК содержит нетранслируемые последовательности между кодирующими последовательностями. Рестрикционные карты позволили уточнить локализацию таких вставочных последовательностей.
Рис. 26.7. Выявление вставочных последовательностей с помощью электронной микроскопии. Молекула мРНК (красная линия) гибридизуется с геномной ДНК, содержащей соответствующий ген. А - если ген непрерывен, видна одна петля одноцепочечной ДНК (показано синим цветом); Б - если ген содержит вставочную последовательность, видны две петли одноцепочечной ДНК (показано синим цветом) и одна петля двухцепочечной ДНК (синий и желтый цвет)
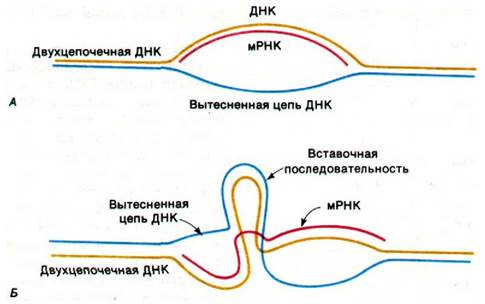
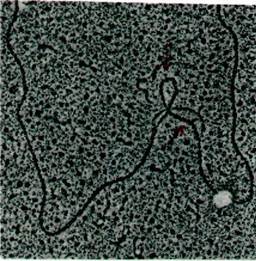
Рис. 26.8. Электронная микрофотография гибрида ß-глобиновой мРНК и фрагмента геномной ДНК, содержащего ген ß-глобина. Толстые петли двухспиральной ДНК - вставочные последовательности в ДНК, которых нет в мРНК (как на рис. 26.7, Б). Верхняя стрелка указывает на большую вставочную последовательность, нижняя стрелка - на маленькую
На каком этапе экспрессии гена удаляются вставочные последовательности? Новосинтезированные РНК, выделенные из ядра, значительно длиннее молекул мРНК, которые из них получаются. В частности, первичный транскрипт гена в-глобина содержит две нетранслируемые области. Эти вставочные последовательности в первичном 15S-транскрипте вырезаются, а кодирующие последовательности одновременно соединяются под действием фермента сплайсинга. Этот фермент обладает высокой точностью. Так, образуется зрелая 9S-мРНК (рис. 26.9). Кодирующие последовательности прерывистых («разорванных») генов называются экзонами, а вставочные последовательности - интронами (от англ. слов expressed regions - экспрессирующиеся участки и intervening sequence - прерывающая последовательность).
Рис. 26.9. Транскрипция гена в-глобина и удаление вставочных последовательностей из первичного РНК-транскрипта. Образование «колпачка» и присоединение poly (A) обсуждаются в разд. 29.22
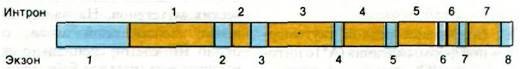
Еще один прерывистый эукариотический ген - ген овальбумина куриного яйца, который состоит из восьми экзонов, разделенных семью длинными интронами (рис. 26.10). Еще более удивителен ген ко- нальбумина: он содержит не менее 17 экзонов. Общее свойство экспрессии этих генов - то, что их экзоны располагаются в мРНК и в ДНК в одной и той же последовательности. Таким образом, прерывистые гены, подобно непрерывным генам, коллинеарны своим полипептидным продуктам.
Рис. 26.10. Структура гена овальбумина куриного яйца. Интроны (некодирующие участки) показаны желтым цветом, а экзоны - синим
Все картированные до настоящего времени гены птиц и млекопитающих, кроме генов гистонов (разд. 29.13), содержат интроны. Почему практически все гены высших эукариот содержат вставочные последовательности? Один из возможных ответов состоит в том, что прерывистые гены отражают процесс эволюции. Экзоны могут соответствовать крупным структурным или функциональным элементам (доменам), которые соединились и образовали белки с новыми свойствами. Другая возможность состоит в том, что вырезание вставочных последовательностей регулирует поток мРНК из ядра в цитозоль. В соответствии с этой гипотезой сплайсинг (сращивание) первичного транскрипта играет ключевую роль: он определяет, какие белки синтезирует клетка. Открытие прерывистых генов у высших организмов ввело нас в увлекательную область познания, которая, по-видимому, имеет большое значение для понимания роста и дифференцировки клеток.