БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 29. ЭУКАРИОТИЧЕСКИЕ ХРОМОСОМЫ И ВЫРАЖЕНИЕ ГЕНОВ У ЭУКАРИОТ
29.3. Последовательности аминокислот в гистонах Н3 и Н4 почти одинаковы у всех животных и растений
Эмиль Смит и Робер Де-Ланж (Emil Smith, Robert DeLange) показали, что последовательности аминокислот в гистоне Н4 из проростков гороха и из тимуса теленка различаются только по двум положениям из 102. Замены эти очень незначительны: валин - вместо изолейцина и лизин - вместо аргинина. Таким образом, последовательность аминокислот гистона Н4 сохранилась почти без изменений на протяжении 1,2 • 109лет, прошедших со времени разделения всего живого на царства растений и животных. Гистон Н3 также мало изменился на протяжении этого колоссального периода эволюции. Последовательности аминокислот в гистоне Н3 из проростков гороха и из тимуса теленка различаются по четырем положениям. Интересно сравнить скорость изменения последовательности этих гистонов в ходе эволюции со скоростью изменения других белков. Обычно для этого используется величина единичного эволюционного периода. Она равна времени, за которое последовательность аминокислот изменяется на 1% после того, как дивергируют две эволюционные линии. Эта величина для гистонов Н3 и Н4 составляет 3 • 108 и 6 • 108 лет соответственно и значительно выше, чем для других исследованных до настоящего времени белков. Например, для цитохрома с единичный эволюционный период составляет 2 • 107 лет, для гемоглобина - 6•106 лет, для фибринопептидов - 1 • 106 лет. Замечательная консервативность структуры гистонов Н3 и Н4 свидетельствует о том, что они выполняют какую-то чрезвычайно важную функцию, возникшую на заре эволюции эукариот и сохранившуюся с тех пор почти без изменений.
Рис. 29.3. Последовательность аминокислот в гистонах Н4 из тимуса теленка. Несколько остатков модифицированы. α-Амино- группа ацетилирована, равно как и е-аминогруппа Lуs-16. е- Аминогруппа Lуs-20 метилирована или диметилирована. Гистон Н4 из проростков гороха имеет такую же последовательность аминокислот, за исключением положений 60 (изолейцин) и 77 (аргинин)

29.4. Нуклеосомы - повторяющиеся субъединицы хроматина
Как происходит взаимодействие гистонов с ДНК и образование нити хроматина? Основываясь на данных, полученных различными методами, Роджер Корнберг (Roger Kornberg) высказал в 1974 г. предположение, что хроматин состоит из повторяющихся субъединиц, каждая из которых включает 200 пар оснований ДНК и по 2 молекулы гистонов Н2А, Н2В, Н3 и Н4. Теперь эти повторяющиеся единицы называют нуклеосомами. Большая часть ДНК намотана на гистоновую сердцевину (гистоновый кор). Остальная ДНК, так называемый линкер, или межнуклеосомная ДНК, соединяет соседние нуклеосомы и обеспечивает гибкость хроматиновой нити. Таким образом, хроматиновая нить представляет собой гибкую цепочку нуклеосом, напоминающую бусины на нитке.
Целый ряд экспериментальных данных свидетельствует в пользу этой модели структуры хроматина.
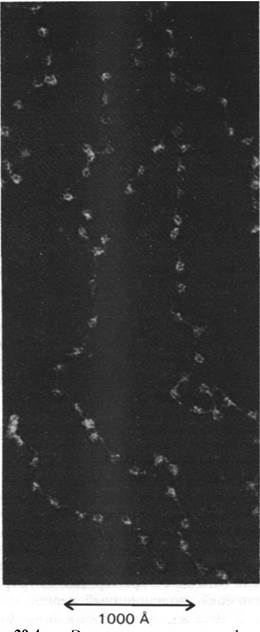
1. Электронная микроскопия. На электронных микрофотографиях хроматина видны группы расположенных друг за другом бусин диаметром 100 А, соединенных тонкой нитью (рис. 29.4). Степень растяжения хроматиновой нити зависит от способа подготовки образцов для микроскопии. Некоторые методы дают электронные микрофотографии с более компактным расположением 100-ангстремных бусин. Следовательно, электронная микроскопия прямо подтверждает, что хроматин - цепочка почти сферических частиц, между которыми расположены гибкие участки.
Рис. 29.4. Электронная микрофотография хроматина. Частицы, похожие на бусины, имеют диаметр около 100 А
2. Дифракция рентгеновских лучей и нейтронов. При рентгеновской дифракции на нитях хроматина также виден повтор длиной 100 А. Нейтронная дифракция показывает, что ДНК расположена снаружи нуклеосомы.
3. Нуклеазный гидролиз. Свободную ДНК в растворе можно расщепить по любой из ее фосфодиэфирных связей с помощью панкреатической дезоксирибонуклеазы I (ДНКазы I) или микрококковой нуклеазы. ДНК в хроматине, за исключением нескольких участков, наоборот, защищена от гидролитического действия нуклеазы. Характер расщепления хроматина поражает своей простотой: на электрофореграмме видна лесенка четко выраженных полос (рис. 29.5). В этих фрагментах содержатся фрагменты ДНК, кратные основному повтору длиной примерно 200 пар оснований. Электронные микрофотографии показывают, что число сферических частиц во фрагменте хроматина равно числу 200-парных повторов (рис. 29.6). Например, фрагмент, содержащий ДНК длиной 600 пар оснований, состоит из трех 100-ангстремных частиц. Следовательно, бусина, видимая на электронной микрофотографии, соответствует нуклеосоме, получаемой при нуклеазном гидролизе.
4. Реконструкция. Если добавить гистоны к ДНК аденовируса или обезьяньего вируса SV-40, можно получить in vitro xpoматиноподобную нить. Количество ДНК, связанной с нуклеосомой в таких системах реконструкции, составляет около 200 пар оснований. Кроме того, для образования нуклеосомы необходимо эквимолярное количество гистонов Н2А, Н2В, Н3 и Н4. Если какого-либо гистона в смеси для реконструкции оказывается недостаточно, образования характерных бусин не происходит. Однако гистон Н1 для реконструкции не нужен; этот факт перекликается с тем, что гистон Н1 присутствует не во всех нуклеосомах эукариотических клеток. Исследования дифракции рентгеновских лучей также показывают, что для получения картины, характерной для хроматина, необходимо добавить гистоны Н2А, Н2В, Н3 и Н4.
Рис. 29.5. Гель-электрофорез фрагментов ДНК хроматина, полученных с помощью ограниченного гидролиза микрококковой нуклеазой. На дорожку А была нанесена нефракционированная смесь. При центрифугировании в градиенте концентрации сахарозы были получены фракции мономеров (Б), димеров (В), тримеров (Г) и тетрамеров
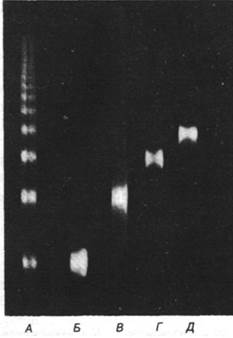
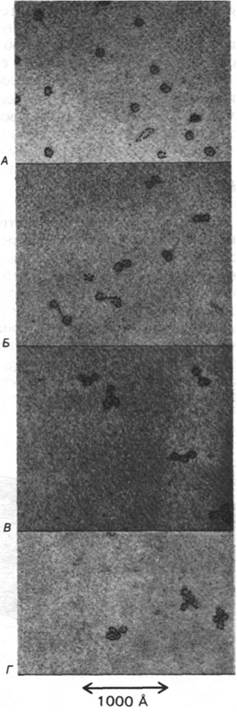
Рис. 29.6. Электронные микрофотографии мономеров (А), димеров (Б), тримеров (В) и тетрамеров (Г) нуклеосом, полученных, как описано в подписи к рис. 29.5
29.5. Минимальная нуклеосома («ядро» нуклеосомы) состоит из ДНК длиной 140 пар оснований, намотанной на октамер гистонов
Содержание ДНК в нуклеосомах различных организмов и типов клеток колеблется от 160 до 240 пар оснований (табл. 29.2). Что лежит в основе этих различий? И в этом случае использование нуклеаз оказалось весьма информативным. Нуклеосомы можно в свою очередь гидролизовать микрококковой нуклеазой; при этом получаются частицы минимальной нуклеосомы (нуклеосомного «ядра»), содержащие 140 пар оснований, независимо от исходного содержания ДНК в расчете на одну нуклеосому. Эта минимальная нуклеосома, по всей вероятности, практически одинакова у всех эукариот. Она состоит из фрагмента ДНК длиной 140 пар оснований, связанного с октамером гистонов (по два гистона Н2А, Н2В, Н3 и Н4).
Таблица 29.2. Содержание ДНК в нуклеосомах

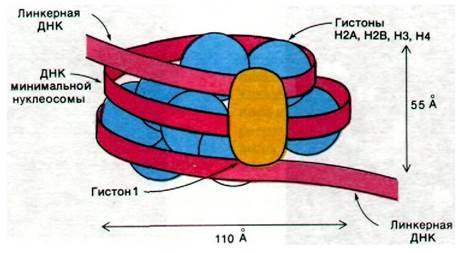
Минимальные нуклеосомы были получены в кристаллическом виде и в настоящее время исследуются с помощью электронной микроскопии (рис. 29.7) и дифракции рентгеновских лучей. Кристаллизация этих частиц показывает, что нуклеосомы хроматина весьма гомогенны. Арон Клуг и Джон Финч (Aaron Klug, John Finch) установили, что минимальная нуклеосома (нуклеосомное «ядро») представляет собой уплощенную частицу размером 110 х 100 x 55 А и состоит из двух слоев. Фрагмент ДНК
длиной 140 пар оснований намотан снаружи на сердцевину и образует 13/4 оборота левозакрученной суперспирали с шагом примерно 28 А (рис. 29.8).
Рис. 29.7. Электронная микрофотография кристалла минимальных нуклеосом. Центры соседних нуклеосом в этом гексагональном слое находятся на расстоянии 100 А друг от друга

Рис. 29.8. Схематическое изображение нуклеосомы. Двойная спираль ДНК (красная полоса) намотана на октамер гистонов (по две молекулы Н2А, Н2В, Н3 и Н4; изображены голубым). Гистон H1 (желтый цвет) связывается с наружной стороной этой минимальной нуклеосомы и с линкерной ДНК
Как уже упоминалось, гистон Н1 не всегда присутствует в нуклеосоме. Последовательность аминокислот в гистоне Н1 наиболее вариабельна из всех пяти гистонов. К тому же гистон Н1 отличается от других гистонов по стехиометрии: на нуклеосому приходится одна молекула гистона Н1. Кроме того, гистон Н1 легко отделяется от нуклеосом, что указывает на его периферическую локализацию, т.е. он не является частью гистоновой сердцевины. Нуклеосомы теряют гистон Н1, когда ДНК в их составе укорачивается со 160 до 140 пар оснований. Таким образом, гистон Н1 почти наверняка расположен вне нуклеосомы, ближе к линкерной ДНК. Гистон Н1 может служить мостиком между различными нуклеосомами и способствовать таким образом повышению компактности хроматина.