БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 30. ВИРУСЫ
30.5. В упорядоченной сборке фага Т4 участвуют вспомогательные белки и протеазы
Сборка фага Т4 - гораздо более сложный процесс, чем сборка ВТМ, так как капсид фага Т4 значительно сложнее по своей структурной организации и содержит примерно 40 различных белков. Еще тринадцать дополнительных белков (по сравнению с ВТМ) участвуют в сборке фага Т4, но не входят в состав капсида. Механизм сборки этого вируса был исследован с помощью сочетания генетических, биохимических и электронно-микроскопических методов. Работы Вильяма Вуда и Роберта Эдгара (William Wood, Robert Edgar), посвященные мутантам фага Т4, дефектным по способности к сборке, позволили установить следующее.
1. Существует три основных пути превращений, которые приводят к образованию вируса. В результате этих превращений независимо формируются головка, отросток и нити отростка (рис. 30.11). Если блокировать образование одного из перечисленных компонентов, это не повлияет на синтез двух других.
Рис. 30.11. Морфогенез фага Т4. Числа возле стрелок обозначают гены, продукты которых необходимы для соответствующих этапов сборки
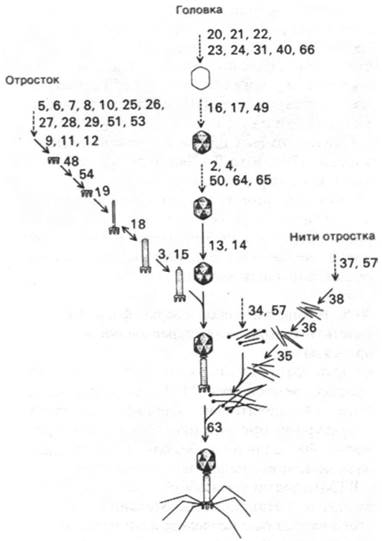
2. Каждая из этих последовательностей превращений идет в строго определенном порядке. Все белки капсида синтезируются одновременно во второй половине цикла заражения. Таким образом, строгой последовательности сборки головки, отростка и нитей отростка способствуют структурные особенности самих промежуточных продуктов. Ни один из этих процессов ассоциации не может идти с заметной скоростью, пока не закончится предыдущий. Возможно, часть энергии связывания на каждом этапе используется для снижения энергии активации следующего процесса ассоциации и таким образом увеличивает его скорость.
3. Головка и отросток должны быть полностью собраны, прежде чем они соединяются друг с другом. Затем готовые нити отростка присоединяются к базальной пластинке. И в этом случае строгая последовательность событий обеспечивает выход только готовых вирусных частиц.
Образование вирионов фага Т4 идет не только путем самосборки. Важную роль на некоторых этапах этого процесса играют вспомогательные (морфопоэтические) белки и протеазы. Например, для образования центральной «втулки» базальной пластинки отростка нужны три белка, которые не входят в состав собранного отростка. Эти вспомогательные белки служат временными шаблонами для ассоциации компонентов «втулки». Протеазы играют важную роль в сборке головки. Основной белок головки с массой 45 кДа, который называется gp23* (gp - от англ. gene product - продукт гена), образуется из предшественника gp23 с массой 55 кДа. Расщепление происходит в тот момент, когда головка частично собрана; это свидетельствует о том, что оно запускает механизм втягивания ДНК в головку. Известны еще три белка головки, которые расщепляются в процессе сборки. Таким образом, фаг Т4 образуется путем самосборки и сборки с участием вспомогательных (морфопоэтических) белков и ферментов.
30.6. В репликации фага T4 участвует конкатемерный промежуточный продукт
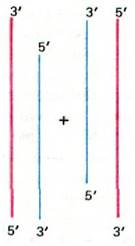
При репликации линейных молекул ДНК, в частности ДНК фага Т4, возникает особая проблема. 5'-концы новообразованной дочерней ДНК застроены не до конца, так как РНК-затравка была удалена, но не была замещена ДНК (рис. 30.12). Напомним, что ДНК-полимераза не способна синтезировать цепи ДНК de novo в направлении 3' → 5' (разд. 24.19). При репликации кольцевых молекул ДНК такой проблемы не возникает, так как 3'-конец новой цепи служит затравкой для завершения синтеза дочерней цепи. Как же решают эту проблему фаг Т4 и другие вирусы, геном которых представляет собой линейную ДНК? Важным указанием на то, как решается эта проблема, послужило открытие, что эти линейные молекулы обладают концевой избыточностью, т.е. последовательность оснований левого конца ДНК в точности повторяется на правом конце:
![]()
Рис. 30.12. 5'-концы новосинтезированных линейных молекул ДНК застроены не до конца. Родительские цепи ДНК показаны красным цветом, а дочерние - синим
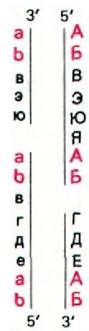
К тому же при репликации этих молекул ДНК образуются длинные конкатемеры. Эти открытия позволили предложить механизм заполнения 5'-концов дочерних цепей. Поскольку одноцепочечные концы новообразованных двухцепочечных молекул взаимно комплементарны, они быстро реассоциируют (рис. 30.13). Эта комплементарность - следствие концевой избыточности последовательности оснований. В конкатемерной цепочке, состоящей из повторяющихся звеньев - двухспиральных молекул фаговой ДНК, 3'-конец одной молекулы служит затравкой для заполнения 5'-конца другой.
Рис. 30.13. Конкатемерный промежуточный продукт репликации линейных двухцепочечных молекул ДНК. Двухцепочечные молекулы ДНК с незастроенными концами ассоциируют друг с другом благодаря спариванию комплементарных одноцепочечных концов (АВ с ab). Затем одноцепочечные пробелы застраиваются
30.7. ДНК фага Т4 вводится в преобразованную головку
Как образуется молекула ДНК, соответствующая одному фаговому геному, и как она упаковывается в головку? Эта проблема невероятно сложна: ДНК имеет контурную длину 56 мкм и при этом должна уместиться в головку, длина которой по большей оси 0,1 мкм. К тому же объем ДНК (1,8 • 10-4 мкм3) ненамного меньше объема головки (2,5 • 10-4 мкм3). A prioriимеются две возможности: ДНК может входить в предобразованную головку или головка может собираться вокруг «ядра» конденсированной ДНК. Выделение пустых головок фага, способных упаковать ДНК, прямо доказывает, что ДНК фага Т4 вводится в предобразованные головки (рис. 30.14). В то время как ДНК входит в головку и свертывается в структуру, напоминающую моток пряжи, происходит расщепление нескольких белков головки.
Рис. 30.14. Схема упаковки ДНК при сборке головки фага Т4
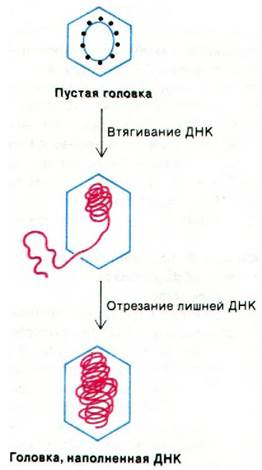
Одновременно происходит разбухание головки. Наконец, когда в головку входит фрагмент ДНК, соответствующий длине одного генома, конкатемерная ДНК расщепляется. Нуклеаза действует не путем узнавания какой-либо определенной последовательности, а расщепляет ДНК именно в тот момент, когда головка наполнена. Этим и объясняется тот факт, что концы молекул ДНК Т4 обладают концевой избыточностью, ДНК упаковывается таким образом, что она может быть очень быстро введена в бактерию при следующем цикле заражения. Чем обеспечивается такая поразительная подвижность, остается загадкой.