БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 30. ВИРУСЫ
30.11. Белки вируса полиомиелита образуются путем множественного расщепления гигантского предшественника
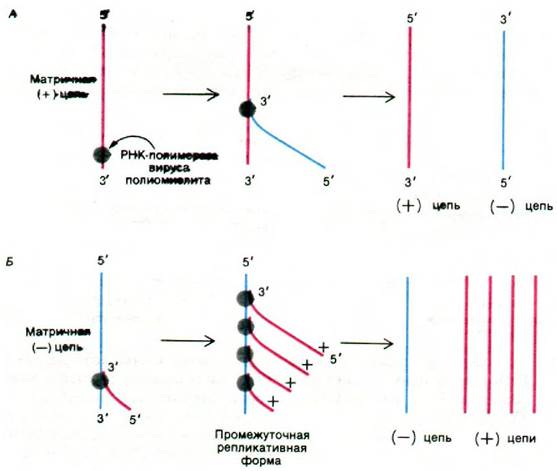
Вирус полиомиелита состоит из одноцепочечной (+) РНК длиной 7,5 kb, заключенной в икосаэдрический капсид. Проникнув в цитоплазму клетки-хозяина, эта молекула РНК вириона используется в качестве матрицы для синтеза белка. Она транслируется рибосомами клетки-хозяина с образованием белков капсида и особой РНК-полимеразы, работающей по РНК-матрице (РНК- репликазы). Затем РНК-репликаза синтезирует (-) цепи на матрице (+) РНК вириона (рис. 30.21). (-) РНК в свою очередь служит матрицей для синтеза множества (+) цепей, которые участвуют в синтезе белка или упаковываются в капсиды и дают новые вирионы.
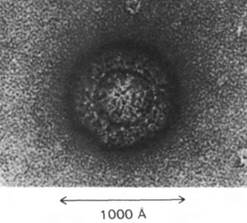
Рис. 30.20. Электронная микрофотография частиц вируса полиомиелита
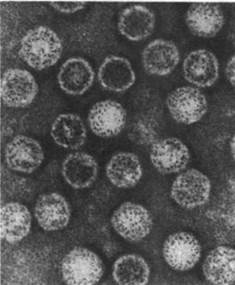
Рис. 30.21. Репликация РНК вируса полиомиелита
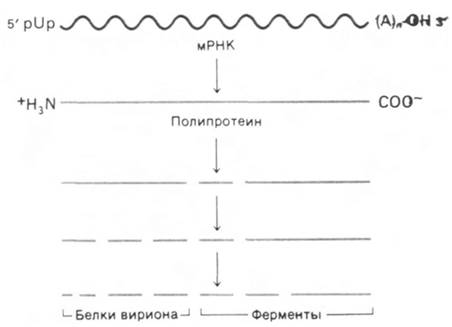
Поразительная особенность выражения генов вируса полиомиелита состоит в том, что (+) РНК вириона служит матрицей для синтеза непрерывной полипептидной цепи, содержащей более 2000 аминокислотных остатков. Дэвид Балтимор (David Baltimore) показал, что этот гигантский полипептид расщепляется протеазами клетки-хозяина на семь белков: четыре белка оболочки, одну РНК-репликазу и два белка, функция которых пока еще неизвестна (рис. 30.22). Новообразованная полипептидная цепь расщепляется на три куска, которые затем подвергаются дальнейшему расщеплению. В частности, два белка оболочки образуются из предшественника на завершающей стадии сборки вириона.
Рис. 30.22. Синтез белков вируса полиомиелита путем множественного расщепления гигантского полипептидного предшественника
Почему вирус полиомиелита синтезирует свои белки таким на первый взгляд сложным путем? По-видимому, у него нет выбора. Напомним, что в эукариотических клетках молекула мРНК по непопятной пока причине может при трансляции дать только одну полипептидную цепь. Прокариотические мРНК, наоборот, сплошь и рядом полицистронны (например, мРНК лактозного оперона). Таким образом, вирус полиомиелита использует расщепление полипротеина, чтобы преодолеть ограничения, накладываемые особенностями животной клетки-хозяина.
30.12. С геномной РНК вируса везикулярною стоматита транскрибируется пять моноцистронных мРНК
Вирус везикулярного стоматита вызывающий легкое заболевание крупного рогатого скота, и вирус бешенства - примеры второго способа выражения генов. Их вирионы содержат одноцепочечную молекулу (-) РНК, которая не может служить матрицей. Поэтому первый этап ее экспрессии - синтез (+) РНК. Поскольку в незараженных клетках нет РНК-репликазы, вирусы должны содержать этот фермент в составе вириона и вводить его при заражении в клетку. Действительно, два из пяти белков вириона ВВС осуществляют репликацию РНК. Для проявления инфекционности вируса необходимы белок L массой 200 кДа (от англ. large - большой) и белок NS массой 45 кДа (от англ. nonstructural - неструктурный), присутствующие в небольших количествах. Геномная РНК находится в комплексе с большим числом молекул белка нуклео- капсида N массой 50 кДа - основного белка вириона. ВВС заключен в липидную двуслойную мембрану, которую он захватывает из плазматической мембраны в процессе отпочковывания от клетки (рис. 30.23). Белок G (от англ. glycoprotein - гликопротеин) массой 65 кДа, кодируемый вирусом, образует шипы, выступающие из этой мембраны. Белок матрикса М массой 29 кДа располагается между оболочкой и нуклео- капсидом. Эти пять белков ВВС образуются при трансляции пяти (+) мРНК, а не путем расщепления одного полипротеина. Та же самая РНК-репликаза синтезирует и длинную (+) РНК, содержащую всю генетическую информацию вируса. Эта полная (+) РНК в свою очередь служит матрицей для синтеза (-) РНК, которая упаковывается и дает новые вирионы.
Рис. 30.23. Электронная микрофотография вируса везикулярного стоматита
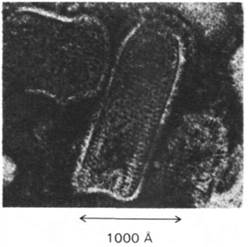
Рис. 30.24. Электронная микрофотография реовируса
Рабдовирусы - вирусы, имеющие форму пули. К ним относятся вирусы везикулярного стоматита и бешенства. Название происходит от греческого слова rhabdo, что означает «палочка».
30.13. Геном реовируса состоит из десяти различных молекул двухцепочечной РНК
Реовирус, содержащий двухцепочечную РНК, поражает клетки млекопитающих и представляет третий тип вирусных генетических систем. Сердцевина вириона содержит десять различных двухцепочечных молекул (±) РНК, ассоциированных с белками. Проникнув в клетку-хозяина, вирион теряет наружную икосаэдрическую оболочку, состоящую из трех видов белков. Удаление этой оболочки активирует РНК-полимеразу, содержащуюся в сердцевине вириона. Эта РНК-зависимая полимераза полностью транскрибирует 10 молекул (+) РНК, так что образующиеся (+) мРНК имеют такую же длину, как фрагменты генома. (±) РНК матрица транскрибируется асимметрично и консервативно, т.е. образуются только (+) РНК, а исходные (±) РНК разрушаются. В сердцевине вириона к 5'-концам этих мРНК присоединяются «колпачки» под действием ферментов. Затем эти концы выходят через каналы в сердцевине (рис. 30.25). Следовательно, сердцевина - высокоорганизованная система синтеза мРНК. Каждая из этих десяти мРНК дает при трансляции один белок. Затем весь набор десяти (+) РНК соединяется с некоторыми вирусными белками и образует предшественник сердцевины («прекор»), в котором синтезируется десять (-) цепей.
Рис. 30.25. Синтез мРНК в сердцевине реовируса. Молекулы мРНК выглядят нитями, отходящими от темных телец-сердцевин
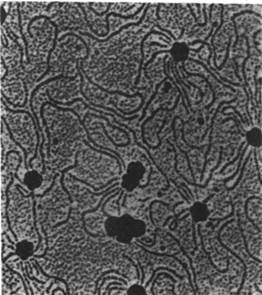
Реовирус - вирус, содержащий двухцепочечную РНК, выделен из дыхательных путей и желудочно-кишечного тракта людей и других млекопитающих; насколько известно, он не вызывает заболевания. Приставка рео составлена из первых букв английских слов respiratory enteric orphan, что означает энтеро-респираторный вирус-сирота (сирота, поскольку бродит «не пристроенный» ни к какой болезни).
Для чего геном реовируса разделен на фрагменты? Как уже было указано выше, вирусы животных не могут иметь полицистронных мРНК. Вирус полиомиелита решает эту проблему путем расщепления гигантского белка-предшественника, а вирус везикулярного стоматита транскрибирует РНК вириона в виде коротких мРНК, каждая из которых соответствует одному белку. Стратегия реовируса состоит в том, чтобы иметь отдельную «хромосому» для каждого синтезируемого белка.
Четвертый путь выражения генетической информации РНК-содержащих вирусов - использование ДНК-посредника, интегрирующей с геном клетки-хозяина. Это более сложная генетическая система, которую используют ретровирусы (РНК-содержащие опухолеродные вирусы). Мы обсудим ее ниже в этой главе (разд. 30.19).
30.14. Мелкие РНК-содержащие фаги содержат перекрывающиеся гены
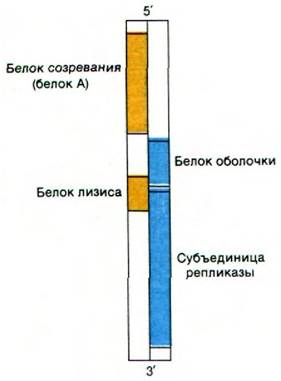
Такие РНК-содержащие фаги, как R17 (MS2, F2) и Qe, относятся к простейшим вирусам. Они имеют форму правильного многогранника и диаметр около 200 А. Капсид этих близкородственных фагов содержит 180 молекул белка оболочки массой 14 кДа и одну молекулу белка А (созревания) массой 38 кДа. Кроме того, одноцепочечная (+) РНК кодирует одну из субъединиц репликазы. До сих пор считалось, что эти мелкие РНК-содержащие вирусы содержат только три гена. Однако обнаружение мутанта фага, образующего нормальные вирионы, но не способного при этом лизировать клетку-хозяина, повлекло за собой поиск еще одного кодируемого вирусом белка. Действительно, у РНК-содержащих фагов имеется четвертый ген, кодирующий белок, необходимый для лизиса бактерии- хозяина. Ген этого белка лизиса перекрывается с генами белка оболочки и субъединицы репликазы (рис. 30.26). Эти мелкие РНК-содержащие фаги, подобно маленькому ДНК-содержащему фагу ɸХ174 (разд. 26.11), используют перекрывающиеся гены для того, чтобы вместить больше информации в свои маленькие геномы. Молекула (+) РНК вириона служит матрицей и для синтеза четырех белков, и для синтеза (-) РНК. Затем (-) РНК используется в качестве матрицы для образования множества копий (+) РНК. Таким образом, по генетической системе эти фаги напоминают вирус полиомиелита.
Рис. 30.26. Перекрывающиеся гены в РНК вирусов R17 и Qβ. Одна рамка считывания показана желтым цветом, другая - синим
Репликаза, синтезирующая (+)- и (-) цепи фаговой РНК, очень интересный фермент. Он проявляет высокую специфичность к гомологичной фаговой РНК. Поэтому молекулы РНК клетки-хозяина не конкурируют с фаговой РНК при репликации. Qβ-репликаза состоит из четырех субъединиц, из которых только одна кодируется фаговой РНК. Три другие субъединицы репликазы - белки клетки-хозяина, которые фаг приспособил для собственных нужд. Два из них - факторы элонгации синтеза белка EF-Tu и EF-Ts, а третий - один из компонентов 30S-субчастицы рибосомы. Так фаг Qβ создает весьма специфичный фермент максимально экономичным способом.
Регуляторные механизмы обеспечивают правильную временную последовательность трансляции и репликации. (+) РНК служит одновременно матрицей для синтеза белка и для синтеза (-) РНК. Было бы нежелательно, чтобы оба процесса происходили одновременно на одной и той же (+) РНК, поскольку рибосомы, движущиеся в направлении 5' → 3', сталкивались бы с репликазой, движущейся в направлении 3' → 5'. Этого не происходит, так как Qβ-репликаза сильно ингибирует связывание рибосом с (+) РНК до тех пор, пока не синтезируется достаточное число молекул (-) РНК.
Четыре фаговых белка синтезируются в различных количествах. Белок оболочки, который синтезируется на протяжении всего периода инфекции, основной продукт трансляции. Одна из причин этого состоит в том. что рибосомы гораздо прочнее связываются с участком инициации соответствующего цистрона, чем с другими участками инициации в (+) РНК. Кроме того, белок оболочки подавляет трансляцию гена репликазы, блокируя его участок инициации. Таким образом, белок оболочки - специфический релрессор трансляции. Белок А транслируется только с незаконченных молекул (+) РНК, так как в полных молекулах РНК его участок инициации блокирован в результате спаривания оснований. Вторичная структура полной молекулы РНК позволяет транслировать лишь небольшое количество белка А.