БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 30. ВИРУСЫ
30.18. Вирусы SV-40 и полиомы могут вызывать продуктивную инфекцию или трансформацию клеток-хозяев
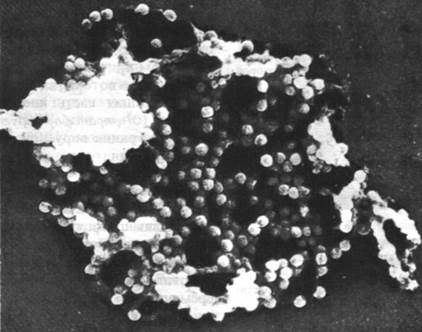
Вирусы SV-40 и полиомы содержат внутри икосаэдрической оболочки маленькую кольцевую двухспиральную ДНК. В некоторых клетках (они называются пермиссивными клетками-хозяевами) развитие этих вирусов идет по пути литического цикла, что приводит к образованию множества новых вирионов (рис. 30.33).
Рис. 30.33. Электронная микрофотография фрагмента ядерной мембраны клетки, зараженной вирусом SV-40. Видны ядерные поры и множество вирионов
При продуктивной инфекции эти вирусы убивают клетки. В клетках других типов (непермиссивных клеток-хозяев) некоторые стадии экспрессии вирусного генома по непонятным причинам блокируются. Никакого вирусного потомства в них не образуется. но небольшая часть клеток - порядка одной на 105 - трансформируется в результате интеграции вирусной ДНК с геномом клетки-хозяина.
К настоящему времени расшифрована полная последовательность 5243 пар оснований ДНК вируса SV-40, а многие аспекты его репликации и транскрипции интенсивно исследуются. Половина ДНК транскрибируется на ранней стадии инфекции, другая половина ДНК - на поздней стадии, одновременно с синтезом вирусной ДНК (рис. 30.34).
Рис. 30.34. Генетическая карта ДНК вируса SV-40, содержащая 5243 пары оснований. Ранняя область (транскрибируется в направлении против часовой стрелки) показана желтым цветом, поздняя область (транскрибируется по часовой стрелке) - синим, место начала репликации (ori) - красным
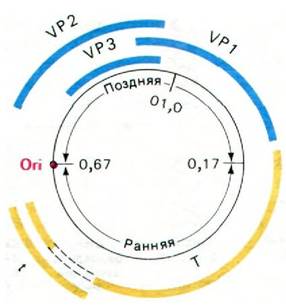
Таблица 30.4. Белки, кодируемые вирусом SV-40
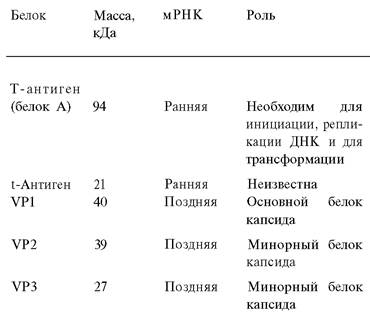
Точка начала репликации находится в той же области, что и начало транскрипции ранней и поздней областей. Ранняя область транскрибируется в направлении против часовой стрелки и кодирует Т-антиген (белок А), необходимый для инициирования репликации ДНК. Другой, иммунологически отличный белок - малый t-антиген - также кодируется ранней областью. При синтезе мРНК для Т-антигена происходит вырезание вставочной последовательности из первичного транскрипта.
Таким образом, вирус SV-40 использует аппарат сплайсинга клеточного ядра. Заслуживает внимания также последовательность оснований в точке начала репликации. Здесь имеются две последовательности длиной по 13 пар оснований, симметричных относительно оси второго порядка, а рядом находится АТ-богатая область:

С этим участком связывается Т-антиген.
Паповавирусы - группа ДНК-содержащих вирусов. Название составлено по названиям трех представителей группы: вирусов папилломы, полиомы и вакуолизирующего вируса (SV-40).
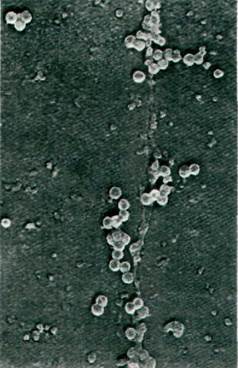
Транскрипция поздней области происходит по направлению часовой стрелки от начала репликации (рис. 30.34) и приводит к синтезу трех белков капсида: VР1, VР2 и VР3. И в этом случае происходит удаление вставочных последовательностей из первичного транскрипта. Три мРНК, по-видимому, образуются в результате различных реакций сплайсинга. N-концевая последовательность аминокислот VР3 перекрывает 70% С-концевой последовательности VР2. Кроме того, перекрывающийся участок из 22 нуклеотидов читается в одной рамке считывания при синтезе VР2 и VР3 и в другой рамке при синтезе VР1. Так, ограниченное количество генетической информации у вируса SV-40 используется с максимальной эффективностью. Еще один пример генетической экономии - то, что SV-40 не синтезирует собственных белков для упаковки ДНК. Новосинтезированная ДНК связывается с гистонами клетки-хозяина (рис. 30.35). Затем этот сверхспирализованный комплекс упаковывается в капсид с помощью белков VР1, VР2 и VР3. В конце концов вирионы потомства высвобождаются при лизисе клетки-хозяина, которая в результате гибнет.
Рис. 30.35. Электронная микрофотография формирующихся частиц опухолеродного вируса SV-40, ассоциированных с клеточной хромосомой
В непермиссивных клетках происходит экспрессия ранней области генома SV-40, поздняя же область не реплицируется и не транскрибируется. Небольшая часть этих клеток трансформируется в результате интеграции генома SV-40 с клеточной ДНК. В отличие от интеграции ДНК фага X в механизме интеграции ДНК SV-40, по-видимому, не участвуют специфические участки ни вирусной, ни клеточной ДНК. Для трансформации, а также, по всей вероятности, для поддержания трансформированного состояния необходимы оба вирусных антигена (Т и t). Введение таких трансформированных клеток чувствительным животным приводит к быстрому образованию опухолей. Основная цель ведущихся в настоящее время исследований вируса SV-40 состоит в том, чтобы выяснить, как экспрессия ранней области интегрированной формы этого вируса делает клетку раковой.
30.19. Ретровирусы содержат обратную транскриптазу, которая синтезирует двухспиральную ДНК, используя в качестве матрицы (+) РНК
Еще один класс опухолеродных вирусов - ретровирусы - содержит (+) РНК-геном в икосаэдрическом «футляре». Это сферическое нуклеопротеиновое «ядро» окружено оболочкой, состоящей из кодируемых вирусом молекул гликопротеина в двуслойной липидной оболочке, происходящей из плазматической мембраны клетки-хозяина. Обычно диаметр ретровирусов составляет 1000 А (см. рис. 30.31).
В 1964 г. Говард Темин (Howard Temin) наблюдал, что заражение такими РНК-содержащими опухолеродными вирусами, как вирус саркомы птиц, блокируется ингибиторами синтеза ДНК. Ингибиторы, например, аметоптерин, 5-фтордезоксиуридин и цитозинарабинозид, эффективны в течение первых двадцати часов после введения вирусов. В результате этого открытия было высказано предположение, что для роста РНК-содержащих опухолеродных вирусов необходим синтез ДНК. К тому же образование частиц вирусного потомства подавляется актиномицином D. Известно, что этот антибиотик ингибирует синтез РНК по ДНК-матрице (разд. 25.18). Так зародилось предположение о том. что для размножения РНК-содержащих опухолеродных вирусов необходима транскрипция ДНК. Эти неожиданные данные привели Темина к мысли, что ДНК-содержащий провирус служит промежуточным продуктом в репликации и в онкогенном действии этих вирусов:

Выдвинутая Темином гипотеза, что генетическая информация может переходить от РНК к ДНК, была вначале холодно встречена большинством исследователей. Она требовала существования неизвестного тогда еще фермента, способного синтезировать ДНК по РНК-матрице (РНК-зависимой ДНК-полимеразы). В 1970 г. Темин и Балтимор (Temin, Baltimore) независимо открыли такой фермент, который называется обратной транскриптазой, в вирионах некоторых РНК-зависимых опухолеродных вирусов. Все вирусы этой группы, исследованные в дальнейшем, содержали обратную транскриптазу, поэтому их и называют ретровирусами (от англ. reverse transcriptase - обратная транскриптаза).
Жизненный цикл обычного ретровируса начинается со связывания вирионов со специфическими рецепторами на поверхности клетки-хозяина и проникновения в клетку. В цитозоле вирусная (+) РНК скидывает оболочку. Затем обратная транскриптаза, содержавшаяся в вирусной частице, синтезирует (-) цепь ДНК. Тот же фермент расщепляет цепь геномной РНК в составе гибрида РНК—ДНК. Теперь обратная транскриптаза синтезирует (+) цепь ДНК, используя в качестве матрицы (-) цепь. Таким образом, обратная транскриптаза осуществляет три последовательные реакции: РНК- зависимый синтез ДНК, гидролиз РНК и ДНК-зависимый синтез ДНК (рис. 30.36).
Рис. 30.36. Синтез ДНК по РНК-матрице обратной транскриптазой. Затравка не показана
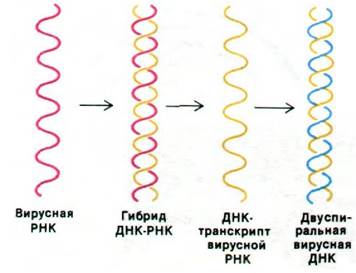
Подобно другим ДНК-полимеразам, обратная транскриптаза синтезирует ДНК в направлении 5' → 3' и неспособна к инициации цепей de novo. Откуда же берется затравка для синтеза вирусной ДНК? Процесс инициации весьма экономичен: (+) РНК вирусного генома содержит нековалентно связанную транспортную РНК (в вирусе саркомы птиц это - триптофановая тРНК), которая была захвачена в клетке-хозяине во время предыдущего цикла заражения. 3 -ОН-группа этой тРНК, основания которой спарены с геномной РНК, действует в качестве затравки синтеза ДНК. Как происходит репликация полной цепи (+) РНК?
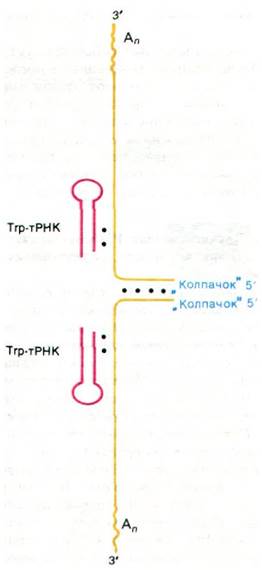
Напомним, что при репликации любой линейной ДНК возникает особая проблема заполнения 5'-концов (разд. 30.6). Ретровирусы нашли очень остроумный выход из этого положения. Их геномы состоят не из одной, а из двух молекул (+) РНК (рис. 30.37). Эти молекулы связаны друг с другом водородными связями вблизи 5'-концов. К тому же (+) РНК содержит одну и ту же последовательность у 5'- и у 3'-конца. Эта концевая избыточность, по- видимому, необходима для процесса репликации, как и в случае фага Т4 (разд. 30.6).
Рис. 30.37. Схематическое изображение генома вируса саркомы птиц. Две идентичные молекулы (+) РНК связаны между собой нековалентно. На 5'-концах находятся «колпачки», а на 3'-концах - poly(А)-последовательности. Молекула тРНК, которая служит затравкой, присоединена посредством спаривания оснований к каждой молекуле РНК
30.20. Ретровирусная ДНК транскрибируется только в том случае, если она интегрирована с геном клетки-хозяина
Двухспиральная вирусная ДНК переходит в кольцевую форму и проникает в ядро. Транскрипция ретровирусной ДНК происходит только после того, как она интегрирует с ДНК клетки-хозяина. Таким образом, в жизненном цикле ретровирусов интеграция — этап обязательный. В жизненном цикле онкогенных ДНК-содержащих вирусов, напротив, интеграция и продуктивная инфекция - альтернативные пути. Еще одно отличие заключается в том, что частота интеграции ретровирусной ДНК очень высока, как и следовало ожидать исходя из ее ключевой роли в продуктивной инфекции.
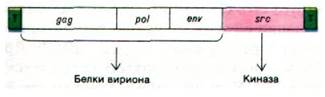
Геном вируса саркомы птиц длиной 10 kb содержит четыре гена (рис. 30.38). Три из них - gag, pol и env - необходимы для продуктивной инфекции. Ген gag кодирует полипротеин массой 76 кДа, который расщепляется на четыре белка, образующих сердцевину вируса. Ген pol кодирует обратную транскриптазу, состоящую из а- и ß- субъединиц. α-Субъединица массой 65 кДа представляет собой фрагмент (массой 90 кДа) протеолиза ß-цепи. Ген env кодирует гликопротеин оболочки вируса, необходимый для прикрепления вируса к поверхности клетки-хозяина. Четвертый ген - src (от англ. sarcomа - саркома) - не нужен для размножения вируса, но необходим для трансформации, как будет показано чуть ниже.
Рис. 30.38. Генетическая карта вируса саркомы птиц. Геном имеет длину 10 kb. Буквой Т обозначены концевые повторяющиеся последовательности
Различные вирусные мРНК, видимо, образуются в результате сплайсинга из первичного транскрипта длиной 10 kb. Затем они транспортируются в цитозоль и здесь транслируются. Геномная РНК и вирусные белки перемещаются к плазматической мембране и включаются в нее. Затем часть измененной мембраны отпочковывается и образует новые вирусные частицы. Таким образом, продуктивная ретровирусная инфекция в отличие от инфекции онкогенными ДНК-содержащими вирусами не является литической. Ретровирусы обычно не убивают клеток-хозяев. Ретровирусная ДНК остается в геноме зараженной клетки и продолжает экспрессироваться. Кроме того, интегрированная вирусная ДНК реплицируется вместе с клеточной ДНК, поэтому дочерние клетки наследуют вирусный геном.