БИОХИМИЯ - Л. Страйер - 1984
ТОМ 3
Часть IV ИНФОРМАЦИЯ
ГЛАВА 31. ПЕРЕСТРОЙКИ ГЕНОВ: РЕКОМБИНАЦИЯ, ТРАНСПОЗИЦИЯ И КЛОНИРОВАНИЕ
31.2. При генетической рекомбинации происходит спаривание гомологичных цепей ДНК с образованием двухцепочечного промежуточного продукта
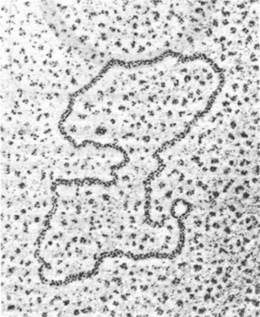
Изображение промежуточных продуктов генетической рекомбинации было получено методом электронной микроскопии. В этих исследованиях была использована ДНК плазмид Е. coli. Как мы вскоре увидим (разд. 31.4), эти небольшие кольцевые двухцепочечные ДНК автономно реплицируются в бактериальной клетке. В присутствии хлорамфеникола число плазмид в одной бактерии увеличивается примерно с 20 до 1000. Этот антибиотик - ингибитор синтеза белка - подавляет репликацию бактериальной хромосомы, но не ингибирует репликации плазмид. Итак, бактериальная клетка оказывается наполненной молекулами плазмид, способными к рекомбинации. Электронная микроскопия плазмид, выделенных из этих клеток, показывает, что примерно четверть из них представляют собой димеры в форме восьмерок (рис. 31.4, А). Затем эти димеры расщепили рестриктирующей эндонуклеазой EcoRI. разрезающей мономер плазмиды в строго определенном месте. Если бы димеры были взаимозацепленными друг с другом мономерами или кольцами двухкратной длины, то под действием рестриктирующей эндонуклеазы образовались бы палочки одинаковой длины. С другой стороны, если два плазмидных кольца ковалентно соединены в области участка гомологии, должна быть видна структура с четырьмя ветвями в форме греческой буквы ӽ. В действительности почти все восьмерки превращаются в %- форму (рис. 31.4, Б). Это служит надежным подтверждением, что восьмерки представляют собой промежуточные продукты репликации. В пользу этого вывода говорит и то, что у некоторых мутантов Е. coli, дефектных по рекомбинации, восьмерки не образуются.
Рис. 31.4. Электронная микрофотография молекул ДНК в процессе рекомбинации: А - промежуточный продукт в форме восьмерки, состоящий из двух молекул ДНК; Б - при расщеплении этого промежуточного продукта рестриктирующей эндонуклеазой образуется х- форма
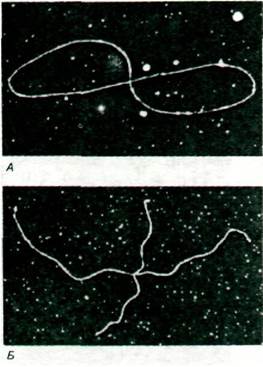
Рис. 31.5. Предполагаемая схема расщепления двух молекул ДНК, соединенных в негомологичных (А) и в гомологичных (Б) участках. Наблюдаемая симметрия Х-форм (как на рис. 31.4, Б) показывает, что молекулы ДНК в форме восьмерок соединены в гомологичных участках
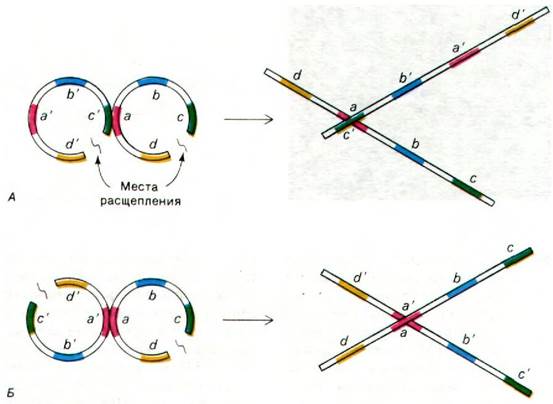
Какова структура области перекрестка двух геномов в восьмерках? Точка контакта (пересечения) х-форм всегда делит всю структуру на две пары плеч равной длины. Это означает, что геномы соединяются в области гомологии (рис. 31.5). Если бы плазмиды соединялись в области негомологичных последовательностей, то длины всех четырех плеч распределялись бы случайным образом. Кроме того, точка контакта располагается примерно с равной вероятностью вдоль всей плазмиды; отсюда следует, что спаривание может происходить во многих положениях. Способ соединения нитей в области перекреста был исследован с помощью его избирательной денатурации. На электронных микрофотографиях видно, что четыре двухцепочечные молекулы в местах соединения двух геномов отходят от кольца, состоящего из одиночных нитей (рис. 31.6). Затем этот промежуточный продукт может быть расщеплен и лигирован, так что образуются две пары различных рекомбинантных молекул (рис. 31.7).
Рис. 31.6. А - электронная микрофотография ӽ-формы; Б - схема молекулы, изображенной на фотографии. Участок гомологии (богатый АТ-парами оснований) был избирательно денатурирован формамидом, чтобы было видно соединение цепей в области перекреста
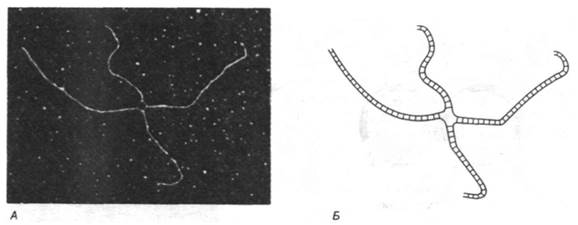
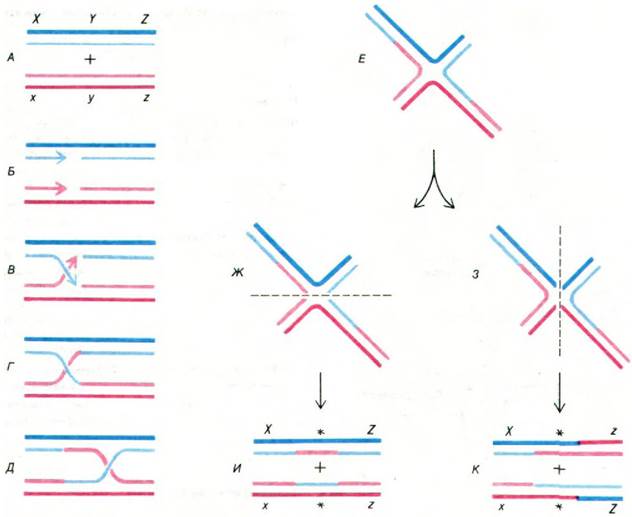
Рис. 31.7. Модель генетической рекомбинации, предложенная Робином Холидеем (Robin Holliday). Одна из родительских двухцепочечных молекул изображена синим цветом, а другая красным. Более темная нить в каждом дуплексе - (+) цепь. Буквами X, Y и Z обозначены три гена; х, у и z - их аллели. В и Г - ковалентное соединение родительских молекул ДНК. Е - иное представление комплекса молекул, изображенного на рис. Д. Обратите внимание, что эта структура на рис. Е может быть разрезана по горизонтальной или по вертикальной оси. Воссоединение нитей на рис. Ж и З дает два различных набора рекомбинантов (И и К). Участки, содержащие по одной цепи каждого из родительских дуплексов, отмечены звездочками
31.3. Белок геcА катализирует АТР-зависимый обмен цепей ДНК при генетической рекомбинации
Процесс, который мы обсуждали до сих пор, называется общей генетической рекомбинацией, поскольку обмены могут происходить между любыми парами гомологичных последовательностей в родительских молекулах ДНК. У Е. coli общая рекомбинация зависит от генов rec. В клетках rec бактериальная ДНК не может рекомбинировать с экзогенными молекулами ДНК. Были идентифицированы три гена rec: recÄ, recВ и recC. Белки recВ (масса 140 кДа) и геg (128 кДа) - две субъединицы одной нуклеазы, которая расплетает двухспиральную ДНК и расщепляет на куски сначала одну из расплетенных цепей, а затем другую. Эти куски, содержащие несколько сотен нуклеотидов, подвергаются дальнейшему расщеплению ферментом recВС, обладающим активностью экзонуклеазы. Расплетающая и нуклеазная активности этого ферментного комплекса зависят от гидролиза АТР. Скорее всего роль белка гесВС в рекомбинации сводится к образованию одноцепочечной ДНК, способной внедриться в двухцепочечную молекулу ДНК.
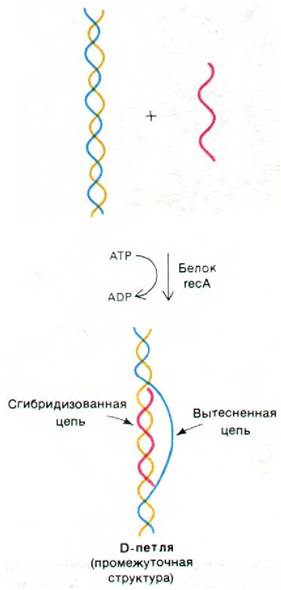
Как одноцепочечная ДНК находит гомологичную последовательность в двухцепочечной молекуле, спаривается с комплементарной цепью и вытесняет вторую цепь? Недавно было показано, что эту реакцию катализирует белок гесА с массой 40 кДа, который гидролизует АТР и использует выделяющуюся энергию. Продукт этой реакции состоит из двухцепочечного участка и вытесненной одноцепочечной петли и имеет форму буквы D; он называется D-петлей (рис. 31.8). Образованию D-петли способствует белок, который специфически связывается с одноцепочечной ДНК. Этот белок играет также важную роль в репликации ДНК (разд. 24.21); он стабилизирует одноцепочечную ДНК, образовавшуюся под действием нуклеазы recВС, и стимулирует внедрение этой цепи в гомологичную двухспиральную молекулу, катализируемую белком гесА.
Рис. 31.8. Спаривание одноцепочечной молекулы ДНК (показана красным цветом) с комплементарной цепью (желтым) дуплекса, катализируемое белком rec A. В результате образуется структура, называемая D-петлей
31.4. Бактерии содержат плазмиды и другие подвижные генетические элементы
Общая генетическая рекомбинация приводит к возникновению новых комбинаций специфических аллелей, но не изменяет расположения локусов. Другими словами, при гомологичной рекомбинации ABCDE с A'B'C'D'E' легко получится ABC'D'E', но не может получиться ABXYZCDE или ABE. Такие крупные генетические перестройки
происходят при участии подвижных генетических элементов (табл. 31.1). Важный класс подвижных генетических элементов - плазмиды. Это кольцевые двухцепочечные молекулы ДНК (рис. 31.9), размер которых колеблется от двух до нескольких сотен тысяч пар оснований (kb). Плазмиды содержат гены, ответственные за инактивирование антибиотиков, метаболизм природных соединений и образование токсинов. В сущности, плазмиды представляют собой дополнительные хромосомы. Они отличаются от бактериальной хромосомы тем, что без них в определенных условиях клетка может обойтись. Кроме того, плазмиды обладают способностью реплицироваться независимо от клеточной хромосомы. Клетка Е. coli обычно содержит около 20 копий мелких хромосом и 1-2 большие.
Таблица 31.1. Мобильные генетические элементы

Рис. 31.9. Электронная микрофотография небольшой плазмиды фактора R