Биохимия в таблицах схемах и графиках - С.Д. Жамсаранова 2009
Трансмембранная передача сигнала
Структура нуклеиновых кислот
Нуклеиновые кислоты - гетерополимеры, их мономерами являются мононуклеотиды.
Роль нуклеиновых кислот
1. ДНК: хранение генетической информации.
2. РНК:
а) хранение генетической информации у некоторых вирусов;
б) реализация генетической информации: м-РНК - матричная РНК, т-РНК (транспортная), р-РНК (рибосомальная);
в) некоторые молекулы РНК способны катализировать реакции гидролиза 3’,5’-фосфодиэфирной связи в самой молекуле РНК. Такие РНК называют рибозимами.
Мононуклеотид состоит из азотистого основания + рибоза у РНК (или дезоксирибоза у ДНК) - вместе они составляют нуклеозид, и остатка фосфорной кислоты.
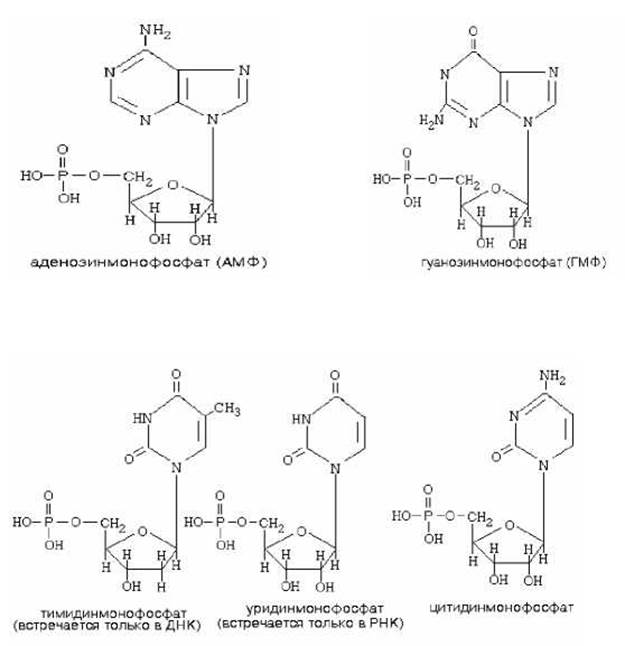
НОМЕНКЛАТУРА НУКЛЕОТИДОВ
Азотистое основание |
Нуклеозид |
Нуклеотид |
аденин |
аденозин |
Аденозинмонофосфат (АМФ) |
гуанин |
гуанозин |
Гуанозинмонофосфат (ГМФ) |
урацил |
уридин |
уридинмонофосфат (УМФ) |
тимин |
тимидин |
тимидинмонофосфат (ТМФ) |
цитозин |
цитидин |
цитидинмонофосфат (ЦМФ) |
В составе нуклеиновых кислот мононуклеотиды связаны 3’,5’-диэфирными связями между рибозами (d-рибозами) соседних мононуклеотидов через остаток фосфорной кислоты.
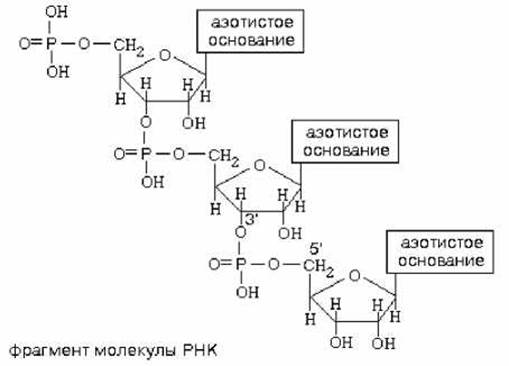
Первичная структура нуклеиновых кислот определяется как последовательность нуклеотидных остатков в полимерной цепи. Как многие другие биополимеры, нуклеиновые кислоты имеют ещё и вторичную структуру, под которой понимают их пространственную организацию.
Вторичная структура ДНК
Молекула ДНК представляет собой правозакрученную спираль, состоящую из двух полинуклеотидных цепей с антипараллельным ходом. Это означает, что 3’-концу одной цепи соответствует 5’-конец другой цепи и наоборот.
СТРУКТУРА ДВОЙНОЙ СПИРАЛИ ДНК
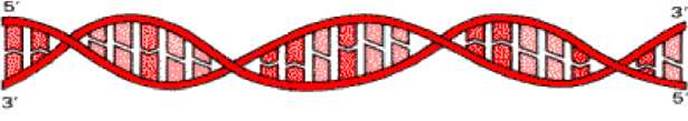
Остатки оснований направлены внутрь спирали. На один виток спирали приходится 10 пар оснований. Цепи ДНК не идентичны, так как нуклеотидный состав их различен, однако первичная структура одной цепи предопределяет нуклеотидную последовательность другой цепи, то есть они комплементарны друг другу. Это связано с существованием комплементарных пар оснований.
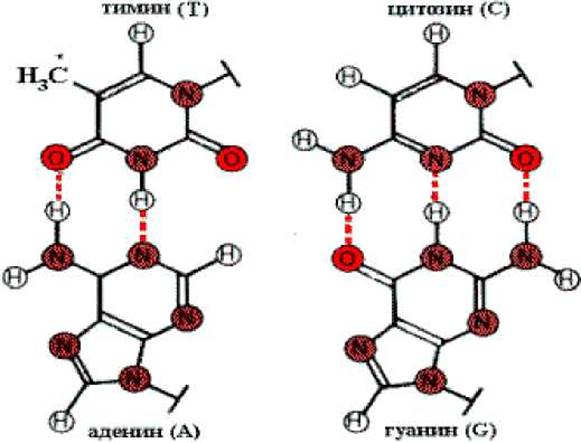
Физико-химическую основу комплементарности составляют водородные связи, которые могут образоваться только между аденином одной цепи и тимином другой, противоположно направленной цепи (пара А-Т), и аналогично между гуанином и цитозином (пара Г-Ц).
Вторичная структура РНК несколько иная. Молекула РНК состоит из одной полинуклеотидной цепи. Отдельные участки этой цепи (до 20-30 нуклеотидных пар) могут быть комплементарны между собой и образовывают спиральную структуру за счет связей между аденином и урацилом (пара A-У) и гуанином и цитозином (пара Г-Ц). Между спирализованными участками располагаются одноцепочечные петли.
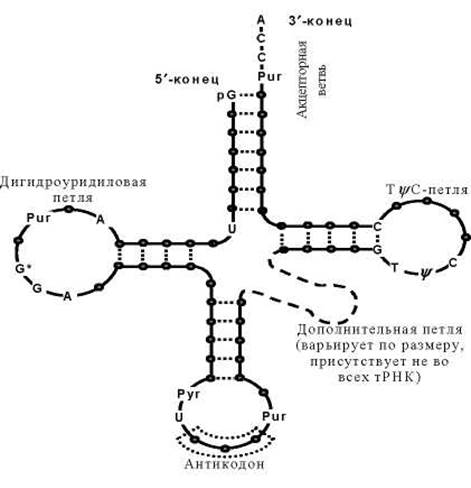
Существует несколько разновидностей РНК: матричная (mRNA), транспортная (тРНК), рибосомная (рРНК). На рисунке приведена структура тРНК, у которой спирализованные участки определяют специфическую пространственную конформацию: фигуру «клеверного листа».
ПРОСТРАНСТВЕННАЯ СТРУКТУРА ТРАНСПОРТНОЙ РНК
«Клеверный лист» — обобщенная вторичная структура, характерная для всех тРНК. Кружками обозначены нуклеотидные остатки, пунктиром — водородные связи между комплементарными основаниями. тРНК имеет на 3’-конце ССА для связывания аминокислоты, а в средней части молекулы - антикодоновый участок - последовательность нуклеотидов, обеспечивающую взаимодействие тРНК с кодоном мРНК.
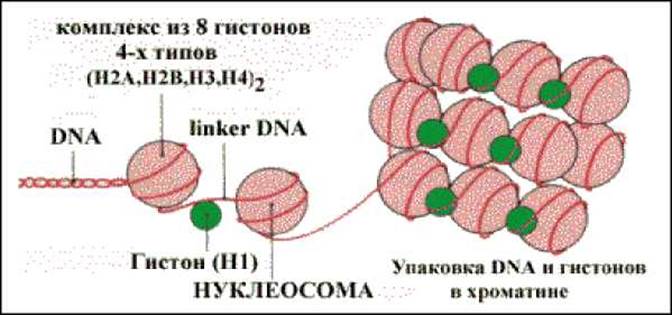
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ХРОМА ТИНА
Каждая молекула ДНК упакована в отдельную хромосому. Хромосомы содержат разнообразные белки, связанные с определенными последовательностями ДНК. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы:
1. Гистоны.
2. Негистоновые белки.
Комплекс белков с ядерной ДНК клеток называют хроматином.
УПАКОВКА ДНК И ГИСТОНОВ С ОБРАЗОВАНИЕМ ХРОМАТИНА
Нуклеиновые кислоты и белки называют информационными молекулами, так как в чередовании их мономеров заложен определенный смысл. Последовательность нуклеотидов в ДНК определяет структуру всех белков клетки. Таким образом, генетическая информация, записанная в ДНК (в генотипе), обеспечивает образование фенотипических признаков клетки, то есть генотип трансформируется в фенотип. Это направление потока информации включает три типа матричных синтезов:
1) синтез ДНК - репликация,
2) синтез РНК - транскрипция,
3) синтез белка - трансляция.
РЕПЛИКАЦИЯ - матричный процесс. Во время репликации каждая из 2 цепей ДНК служит матрицей для образования новой цепи.
Субстраты. Субстратами и источниками энергии для синтеза ДНК явялются дезоксирибонуклеозидтрифосфаты - дНТФ (дАТФ, дГТФ, дТТФ, дЦТФ).
Основные этапы процесса:
1) формирование репликативной вилки;
2) синтез новой цепи ДНК;
3) исключение праймеров. Завершение формирования отстающей цепи ДНК.
Формирование репликативной вилки идет при участии: ДНК-топоизомеразы, которая разрывает цепь (3,5-фосфодиэфирную связь) ДНК, а по окончании репликации зашивает временные надрезы; ДНК-хеликазы, использующей энергию АТФ для расплетания двойной спирали ДНК; белков, дестабилизирующих спираль (или SSB-белков single strand binding). SSB-белки, не закрывая оснований, связываются с одноцепочечной ДНК и этим предотвращают образование «шпилек» и комплементарное скручивание матричных цепей.
ДНК-полимераза δ не способна инициировать синтез новых цепей ДНК, она может лишь удлинять уже имеющуюся нуклеотидную цепь - затравку (праймер). Роль затравки выполняет РНК, синтезируемая специальным ферментом ДНК-полимеразой а.
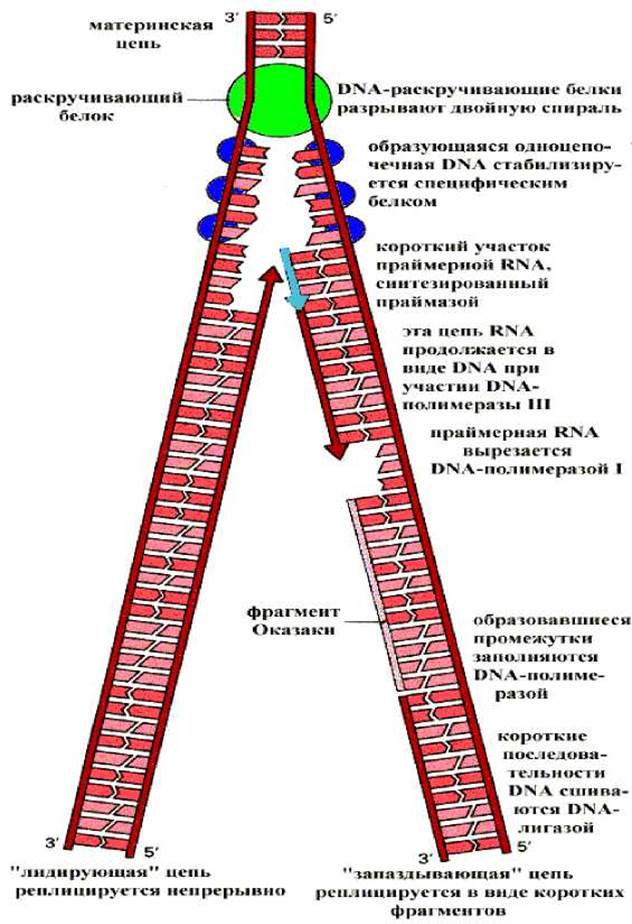
По завершении репликации образуются 2 молекулы двухспиральной ДНК, каждая из которых содержит одну материнскую и одну дочернюю, вновь синтезированную нить. Таким образом репликация обеспечивает воспроизведение генотипа в новых поколениях.