БІОХІМІЯ ДЛЯ ВЧИТЕЛЯ - Ф.Ф. БОЄЧКО - 1985
ОСНОВНІ ХІМІЧНІ КОМПОНЕНТИ КЛІТИН
НУКЛЕЇНОВІ КИСЛОТИ — ДИРИГЕНТИ СИНТЕЗУ БІЛКА
Нуклеїнові кислоти було відкрито понад 100 років тому в ядрах лейкоцитів. Проводячи біохімічні дослідження, виявили в складі їх ядер (nucleus) невідому на той час речовину, яку назвали нуклеїном. Нуклеїн мав властивості кислоти і містив близько 10 % фосфору, що й дало підставу називати його нуклеїновою кислотою.
Згодом нуклеїнові кислоти було виявлено в ядрах і цитоплазмі всіх клітин організму. Проте тривалий час їх розглядали як інертні компоненти. І лише наприкінці першої половини XX ст. погляди на роль цих речовин в організмі змінилися. Нині встановлено, що нуклеїнові кислоти відіграють надзвичайно важливу роль — передають спадкові ознаки та забезпечують синтез білка в організмі.
Хімічний склад нуклеїнових кислот
Нуклеїнові кислоти є полінуклеотидами. їх гігантські молекули складаються з великої кількості структурних одиниць — мононуклеотидів. Останні складаються з пуринових або піримідинових основ, вуглеводних компонентів — пентоз і фосфорної кислоти.
З основ пуринового ряду до складу мононуклеотидів входить в основному аденін (6-амінопурин) і гуанін (2-аміно- 6-оксипурин), які мають плоску структуру з невеликою складкою. Вони є похідними пурину. Гуанін може існувати в двох формах — енольній і кетонній.
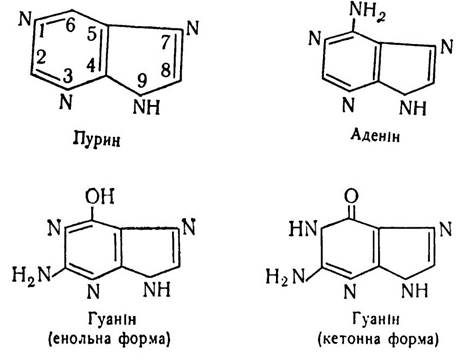
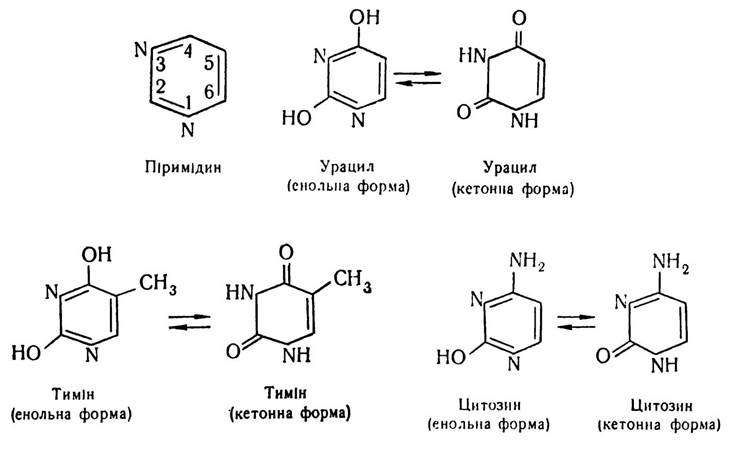
З основ піримідинового ряду до складу мононуклеотидів входять урацил (2,4-діоксипіримідин), тимін (2,4-діоксі-5-метилпіримідин) і цитозин (2-оксі-4-амінопіримідин). Піримідинові основи мають плоску структуру. Вони також можуть перебувати в енольній і кетонній формах.
Крім цих азотистих основ, до складу нуклеїнових кислот у невеликих кількостях (0,1—2 %) входять так звані мінорні (рідкісні) основи: 1-метиладенін, 1-метилгуанін, 5-метил-цитозин, 5-оксиметилцитозин та ін.
Дещо в більших кількостях мінорні основи знайдено в транспортних рибонуклеїнових кислотах. Іноді вміст їх може становити до 20 %.
З групи пентоз до складу мононуклеотидів входять β-D-рибоза і β-D-2-дезоксирибоза. Обидві пентози перебувають в β-D-рибофуранозній формі:

Не так давно було встановлено, що, крім рибози і дезоксирибози, в деяких нуклеїнових кислотах, зокрема в дезоксирибонуклеїновій, може міститись глюкоза.
Третім компонентом нуклеотидів є фосфорна кислота.
Слід зазначити, що пуринові або піримідинові основи, рибоза або дезоксирибоза і фосфорна кислота зв’язані між собою в мононуклеотидах однотипно.
Елементарними ланками нуклеїнових кислот є нуклеотиди, а простішими за них сполуками — нуклеозиди.
Нуклеозиди — двокомпонентні сполуки, побудовані з пуринових або піримідинових основ і пентоз (рибози чи дезоксирибози). Назва їх походить від назви азотистої основи. Так, сполуку аденіну з рибозою називають аденозином, цитозину з рибозою — цитидином, з дезоксирибозою відповідно — дезоксиаденозином і дезоксицитидином.
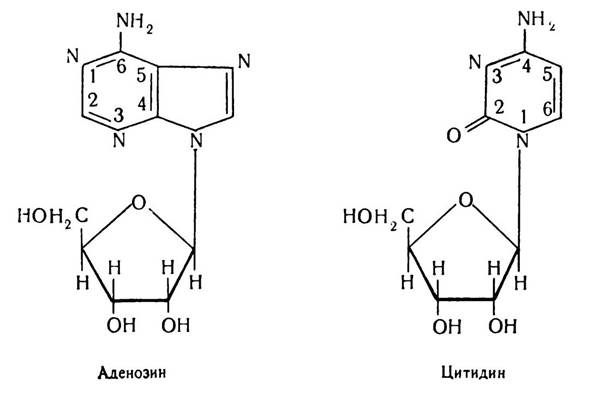
Нуклеозиди, сполучаючись з фосфорною кислотою, утворюють основну структурну одиницю нуклеїнових кислот — нуклеотид. Отже, нуклеотиди є вже трикомпонентними сполуками. До іх складу входять пуринові або піримідинові основи, рибоза або дезоксирибоза і фосфорна кислота.
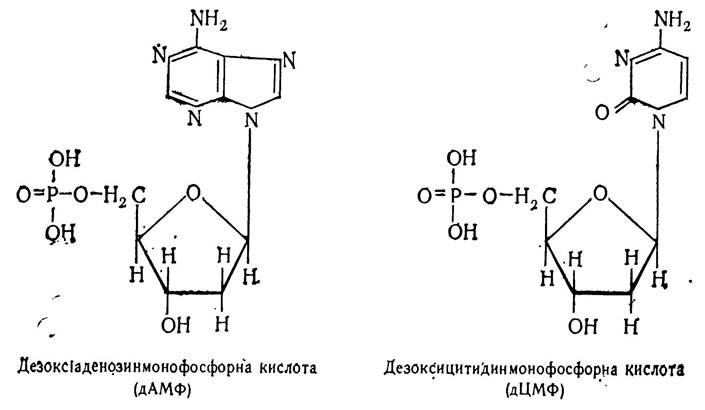
Назва нуклеотидів походить від назви основ, що входять до їх складу, або від назви нуклеозиду. Наприклад, якщо нуклеотид містить азотисту основу аденін, то він називатиметься аденіловою кислотою, або аденозинмонофосфорною (АМФ). Якщо азотиста основа — цитозин, то нуклеотид називатиметься цитидиловою кислотою, або цитидинмонофос форною кислотою (ЦМФ):
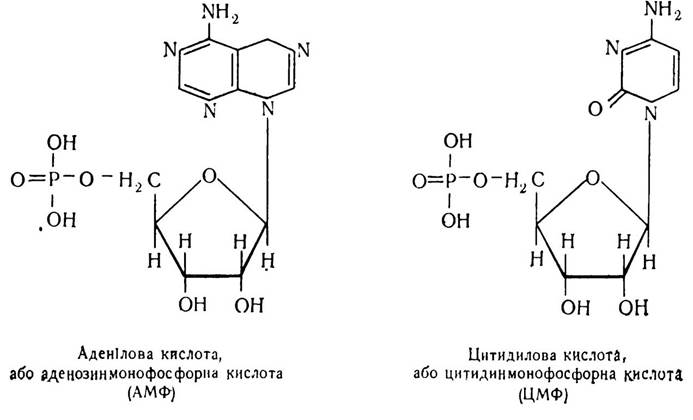
При наявності в нуклеотидах азотистих основ гуаніну або урацилу вони називатимуться відповідно гуаніловою кислотою, або гуанозинмонофосфорною (ГМФ) та уридило- вою кислотою, або уридинмонофосфорною кислотою (УМФ).
Аналогічно називають нуклеотиди, які замість рибози містять дезоксирибозу. Відмінність лише в тому, що до назви нуклеотиду добавляється префікс — дезокси (д):
Нуклеотиди, до складу яких входить рибоза, називають рибонуклеотидами, а якщо входить дезоксирибоза — дезоксирибонуклеотидами.
Встановлено, що нуклеотиди в організмі входять не тільки до складу нуклеїнових кислот, а можуть бути й у вільному стані або складовими частинами ферментних систем. З цього погляду значний інтерес викликає система аденозинфосфорних кислот — АМФ, АДФ і АТФ:
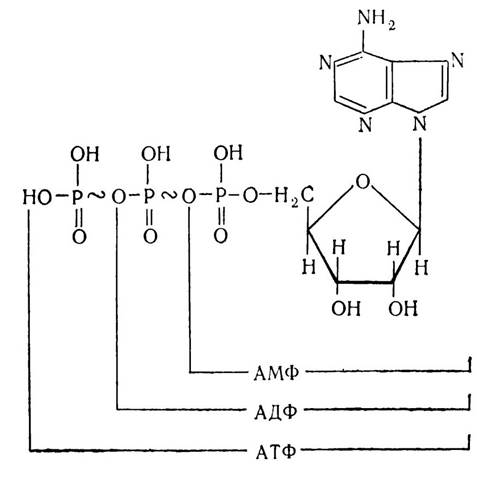
Аденозинмонофосфорна кислота, приєднуючи до свого залишку фосфорної кислоти ще один або два таких залишки, утворює аденозиндифосфорну кислоту (АДФ) або аденозинтрифосфорну кислоту (АТФ). Ці аденозинфосфорні кислоти відіграють важливу роль в обмінних процесах організму. Зокрема, АТФ бере участь в енергетичному обміні організму і є однією з основних макроергічних сполук. Якщо від АТФ відщепити одну або дві молекули фосфорної кислоти, які зв’язані між собою макроергічним зв’язком (~), то виділиться 32,8—42,0 кДж/моль енергії, тоді як енергія звичайного фосфорного зв’язку становить 8— 12 кДж/моль.
Таблиця 7. Компоненти нуклеїнових кислот та їх скорочене позначення
Азотисті основи |
Нуклеозити |
Рибонуклеозидфосфати |
Дезоксирибонуклеози фосфати |
||||
моно- |
ди- |
три- |
моно- |
ди- |
три- |
||
Аденін (А) Гуанін (Г) Цитозин (Ц) Тимін (Т) Урацил (У) |
Аденозин Гуанозин Цитидин Тимідин Уридин |
АМФ ГМФ ЦМФ - УМФ |
АДФ ГДФ ЦДФ - УДФ |
АТФ ГТФ ЦТФ - УТФ |
дАМФ дГМФ дЦМФ дТМФ - |
дАДФ дГДФ дЦДФ дТДФ - |
дАТФ дГТФ дЦТФ дТТФ\- |
В обміні речовин та енергії беруть участь і інші фосфорильовані нуклеозиди, зокрема ті, які містять гуанін, цитозин і урацил. Проте АТФ в процесах обміну речовин та енергії займає основне місце.
Аденозинтрифосфорна кислота внаслідок каталічної дії особливого ферменту — аденілатциклази може відщеплювати два залишки фосфорної кислоти і перетворюватись при цьому в аденозин-3',5'-монофосфорну кислоту, тобто циклічну АМФ:
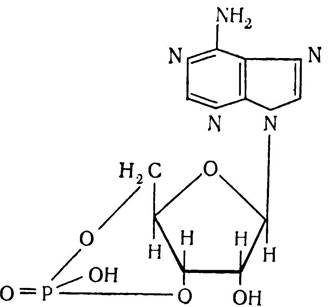
Циклічна АМФ відіграє важливу роль в регуляції протікання ферментативних процесів і в синтезі білків в організмі.
Будова нуклеїнових кислот
Нуклеотиди, які побудовані з пуринових або пїримідинових основ, рибози або дезоксирибози і залишку фосфорної кислоти, сполучаючись між собою, утворюють ди-, три-, тетра-, пента-, гекса- і полінуклеотиди, тобто нуклеїнові кислоти. До складу нуклеїнових кислот входять сотні і тисячі окремих нуклеотндів. Вони з’єднані між собою за допомогою фосфодіефірного зв’язку, який утворюється внаслідок взаємодії гідроксильної групи пентози одного нуклеотиду із залишком фосфорної кислоти пентози наступного нуклеотиду.
Нуклеїнові кислоти, залежно від хімічного складу, структури і біологічної ролі поділяють на дві великі групи: рибонуклеїнові кислоти (РНК) і дезоксирибонуклеїнові кислоти (ДНК).
Рибонуклеїнові кислоти побудовані з рибонуклеотидів, які, крім залишку фосфорної кислоти, містять вуглеводний компонент рибозу і азотисті основи — аденін, гуанін, урацил і цитозин. До складу дезоксирибонуклеотидів входять нуклеотиди, у яких вуглеводний компонент — дезоксирибоза, та азотисті основи — аденін, гуанін, цитозин, а замість урацилу — тимін. Отже, РНК і ДНК відрізняється між собою тим, що перша містить рибозу і урацил, а друга — дезоксирибозу і тимін.
ДНК — основний генетичний матеріал живих систем. В організмах, за винятком вірусів і бактерій, вони зосереджені головним чином в ядрах клітин. Невелика кількість ДНК міститься в мітохондріях, хлоропластах та в деяких включеннях клітин.
Характерною ознакою ДНК є велика відносна молекулярна маса, що коливається в досить широких межах і залежить насамперед від того, з якого організму вона виділена. Так, відносна молекулярна маса очищених препаратів ДНК вірусу поліоми становить 3·106, кишкової палички — 2,5·109, а ДНК людини 3,2·1010—1,6·1011.
Для ДНК, як і для білків, властиві кілька рівнів структур: первинна, вторинна і третинна.
Первинна структура характеризується певною послідовністю розміщення нуклеотидів у полінуклеотидних ланцюгах ДНК. Вивчити цю структуру важко, оскільки різні види ДНК побудовані з великої кількості нуклеотидів.
Крім того, порядок розміщення нуклеотидів у полінуклеотидах для різних видів ДНК різний.
Дослідження хімічного складу нуклеїнових кислот, проведені за останні десятиріччя, показали, що ДНК різних ви* дів організмів відрізняються одна від одної і характеризуються своїм особливим нуклеотидним складом. Водночас були встановлені і певні закономірності нуклеотидних співвідношень, що лежать в основі різних видів ДНК.
На основі таких даних американський учений Е. Чаргафф вивів ряд правил щодо нуклеотидного складу ДНК:
1. Сума пуринових нуклеотидів дорівнює сумі піримідинових нуклеотидів (Пур. = Пір. або -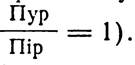
2. Вміст аденіну (А) дорівнює вмісту тиміну (Т) (А = Т, або![]()
3. Вміст гуаніну (Г) дорівнює вмісту цитозину (Ц) (Г = = Ц, або 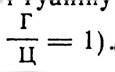
4. Кількість амінних груп біля шостого вуглецевого атома азотистих основ дорівнює кількості кетонних ГОУП у цьому ж положенні: (А+Ц=Г+Т, тобто![]()
5. В одних видах ДНК, зокрема, виділених з організму тварин, вищих рослин і багатьох мікроорганізмів, нуклео- тиди, що містять аденін і тимін, переважають над нуклеотидами, що містять гуанін і цитозин (А+Т>Г+Ц). Такі дезоксирибонуклеїнові кислоти називають ДНК АТ-типу.
В інших ДНК, зокрема, виділених з мікроорганізмів і бактерій, нуклеотиди, які містять гуанін і цитозин, переважають над нуклеотидами, які містять аденін і тимін Г+Ц> >А+Т. Такі ДНК утворюють ГЦ-тип дезоксирибонуклеїнових кислот. У природі переважають ДНК АТ-типу.
Значний вклад у вивчення хімічного складу нуклеїнових кислот внесли радянські вчені А. М. Білозерський і О. С. Спірін. Добуті ними дані дали можливість виявити видову специфічність ДНК у широких масштабах рослинного і тваринного світу.
Вивчення нуклеотидного складу ДНК в різних організмів показало, що він помітно коливається у водоростей, грибів і особливо бактерій. Специфічний склад ДНК в них настільки явно виражений, що може бути однією з надійних систематичних ознак. Нуклеотидний склад ДНК у тварин і вищих рослин, на відміну від мікроорганізмів коливається значно менше. Так, якщо у бактерій коефіцієнт специфічності ДНК, тобто відношення![]() , змінюється в межах від 0,45 до 2,80 (у шість разів), то у вищих рослин і різних видів тварин він перебуває в межах 0,54—0,94 (змінюється лише в два рази).
, змінюється в межах від 0,45 до 2,80 (у шість разів), то у вищих рослин і різних видів тварин він перебуває в межах 0,54—0,94 (змінюється лише в два рази).
Дослідження первинної структури ДНК показало, що в полінуклеотидних ланцюгах відбувається:
1) повторення окремих нуклеотидних блоків
ЦТТЦГА ЦТТЦГА;
ГААГЦТ ГААГЦТ
2) переривання блоків, які повторюються, іншими унікальними блоками:
ЦТТЦГА АТГЦА ЦТТЦГА;
ГААГЦТ ТАЦГТ ГААГЦТ
3) повторення блоків нуклеотидів у зворотному порядку (зворотні повтори):
ЦТТЦГА ТЦГААГ
ГААГЦТ АГЦТТГ.
Блоки, які повторюються, можуть мати різну кількість нуклеотидних залишків.
Оскільки полінуклеотиди ДНК побудовані з великої кількості залишків нуклеотидів, то вивчення їх повної нуклеотидної послідовності становить значні труднощі. Однак уже проведена значна робота щодо успішного розв’язання цього питання. Так, в 1976 р. американський дослідник Г. Корана разом зі своїми співробітниками завершив синтез біологічно активного гена — ділянки дволанцюгових ДНК бактерій кишкової палички, який несе повну генетичну інформацію про біосинтез транспортної тирозинової РНК (тРНКтир). Група радянських і американських дослідників розшифрувала великий фрагмент (283 нуклеотидних залишки) дволанцюгової ДНК дріжджів (ген 5S-pPHK).
Вторинна структура характеризується просторовою конфігурацією полінуклеотидних ланцюгів ДНК- Модель структури молекули ДНК вперше була запропонована вченими із Кембріджського університету в Англії Д. Уотсоном і Ф. Кріком в 1953 р. Відповідно до цієї моделі молекула ДНК являє собою подвійну спіраль, тобто вона складається з двох полінуклеотидних ланцюгів, які закручені правильними витками навколо однієї спільної осі (мал. 8). Без розкручування вони не можуть розійтись між собою. Полінуклеотидні ланцюги ДНК розміщені антипа- ралельно: 5'-кінець одного полінуклеотидного ланцюга міститься проти 3'-кінця другого ланцюга. Вони обернені один до одного азотистими основами, а назовні розміщені залишки дезоксирибози. Діаметр спіралі ДНК становить 2 нм, відстань між спіралями 3,4 нм, кожний виток спіралі містить 10 пар нуклеотидів, отже, кожна пара їх займає 0,34 нм по осі спіралі.
Мал. 8. Схематичне зображення подвійної спіралі ДНК:
1 — за Уотсоном і Кріком; 2 — А-форма ДНК; 3 — В-форма ДНК; с — залишок дезоксирибози; р — залишок фосфорної кислоти.
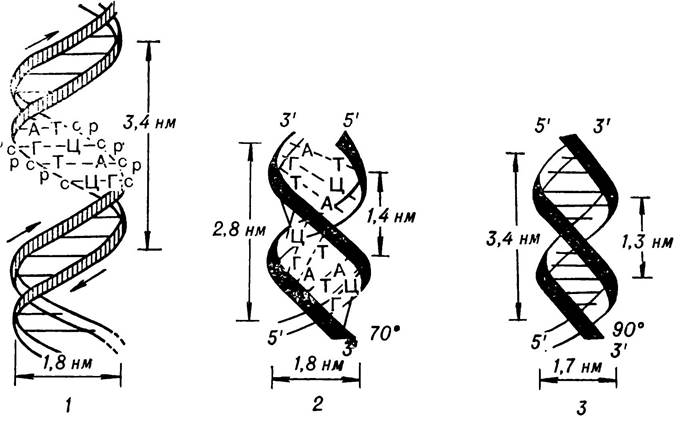
Стабілізація подвійної спіралі відбувається за рахунок водневих зв’язків і гідрофобних взаємодій. Утворення водневих зв’язків у молекулі ДНК — процес точно визначений. Так, аденін одного полінуклеотидного ланцюга завжди зв’язується двома водневими зв’язками з тиміном другого полінуклеотидного ланцюга, а гуанін постійно зв’язується трьома водневими зв’язками з цитозином. Така закономірність дістала назву комплементарності (доповнюваності). У подвійній спіралі ДНК аденін ніби доповнює тимін, а гуанін — цитозин і навпаки. Отже, кожна пара нуклеотидів складається з однієї пуринової основи та однієї піримідинової, які доповнюють одна одну. З такого принципу будови ДНК випливає правило Чаргаффа, за якими вміст аденіну дорівнює вмісту тиміну, а вміст гуаніну дорівнює вмісту цитозину. Важливе значення в стабілізації спіралі ДНК мають неполярні взаємодії з участю делокалізованих π-електронів азотистих основ. Квантово-механічні розрахунки електронної структури ДНК показали, що між комплементарними парами А—Т і Г—Ц відбуваються перекривання π-орбіталей, а це призводить до того, що комплементарні пари азотистих основ утворюють єдину електронну структуру (мал. 9).
Мал. 9. Перекривання я-орбіталей нуклеотидних основ в молекулі
ДНК (за М. Ю. Кеда і Б. І. Кеда).
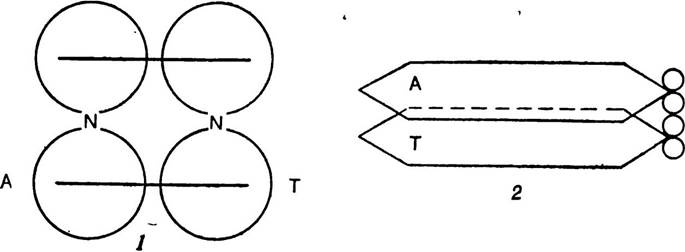
Крім того, перекривання π-електронних систем азотистих основ відбувається і внаслідок їх паралель» ного розміщення всередині подвійної спіралі молекули ДНК (мал. 10). Найбільш значне перекривання я-орбіталей спостерігається тоді, коли пуринові основи містяться під піри- мідиновими. При зворотному порядку розміщення основ перекривання я-орбіталей найменше. Така π-електронна взаємодія між паралельно розміщеними основами або парами основ у молекулі ДНК (так звана стекінг-взаємодія) становить єдину π-електронну систему, яка стабілізує по двійну спіраль ДНК навіть більше, ніж водневі зв’язки.
Двоспіральна ДНК залежно від умов (вмісту води, іонної сили) може набирати певної конформації (мал. 8). Так, на основі вивчення солей ДНК різних лужних металів з різним ступенем вологості виявлено три форми двоспіральної ДНК — А, В і С. Із зміною вологості і виду катіона солі ці форми ДНК можуть переходити одна в одну. В-форма відповідно до моделі Уотсона — Кріка є формою ДНК, яка найчастіше зустрічається в живих організмах. В-форма перетворюється в А-форму, коли вологість препаратів в ДНК становить нижче 70 %. A-форма відрізняється від В-форми тим, що пари азотистих основ розміщені не перпендикулярно до осі спіралі, а під кутом 70°. Внаслідок цього крок спіралі зменшується з 3,4 до 2,8 нм. В A-формі ДНК на один виток припадає одинадцять пар основ (у В-формі— 10), що зумовлює скорочення полінуклеотидного ланцюга приблизно на 25 %. А-конформація часто спостерігається в гібридних молекулах ДНК — РНК, оскільки додаткова гідроксильна група біля другого вуглецевого залишку рибози заважає утворенню В-конформації ДНК.
Мал. 10. Схема π-електронної взаємодії між паралельно розміщеними парами основ в молекулі ДНК (за М. Ю. Кеда і Б. І. Кеда).
С-форма характеризується рихлою і розкрученою структурою. На один виток припадає лише 9 нуклеотидів. Допускають, що в С-формі ДНК міститься в складі хроматину.
Аналіз даних щодо різних форм ДНК свідчить про те, що в біологічному аспекті В-форма найбільш адекватна для процесів реплікації, А-форма — для процесу транскрипції і С-форма — для упаковки ДНК в складі надмолекулярних структур хроматину і деяких вірусів. Отже, вторинна структура молекул ДНК, очевидно, пов’язана із здійсненням інформаційних процесів у живих системах. Так, А-форма — з передачею інформації від ДНК до РНК, В-форма — із примноженням, збільшенням кількості інформації і С-форма — з її збереженням.
Виділення ДНК з різних видів організмів та вивчення її структури показало, що дволанцюгові ДНК окремих прокаріотів — вірусів, бактерій і клітинних органел — мітохондрій та хлоропластів — не лінійні, а замкнуті в кільце, Наприклад, ДНК фага, який називають фагом лямбда, має лінійну форму доти, поки міститься у фаговій частинці. А коли потрапляє в бактеріальну клітину, замикається в кільце, завдяки наявності в ланцюгах ДНК так званих липких кінців. На кожному кінці ДНК один з ланцюгів довший від другого і містить певну кількість неспарених основ. Такі ділянки неспарених основ на обох кінцях молекули комплементарні одна до одної. Тому, коли їх спарюють, кінці ланцюгів ніби злипаються і молекула набуває структури кільця, яким завершується утворення спеціального ферменту (мал. 11). Одноланцюгові кільцеві форми ДНК виявлені у фагах — фх 174, М 13 і S 13 кишкової палички.
Мал. 11. Схема утворення кільцевої форми ДНК фага лямбда.
Водночас слід зазначити, що в ряді мікроорганізмів виявлено одноланцюгові ДНК, які мають як лінійну, так і кільцеву структури. Так, в окремих вірусів (вірусі поліомієліту, дрібному вірусі мишей) знайдено лінійні одноланцюгові ДНК. Вони, на відміну від дволанцюгових ДНК, мають меншу відносну молекулярну масу, часто не зберігається в них відповідне співвідношення між азотистими основами А — Т і Г — Ц.
Третинна структура ДНК характеризується певним розміщенням вторинних структур полінуклеотидних ланцюгів ДНК. Вважають, що характерною особливістю третинної структури ДНК в її спіралізація другого порядку, тобто суперспіралізація. Про це свідчить той факт, що окремі молекули ДНК мають значно меншу довжину, ніж їх повністю витягнуті біспіральні молекули. Наприклад, молекула ДНК з відносною молекулярною масою 10 млн. повинна мати довжину 5 мкм, а насправді її довжина становить лише 0,5 мкм.
Є дані, які свідчать, що ДНК в суперспіралізованому стані міститься в хроматині і хромосомах, причому тут реалізується кілька рівнів суперспіралізації. Так, в ізольованому хроматині окремі ділянки подвійної спіралі обвиваються навколо молекул гістонів і утворюють суперспіраль першого порядку. Комплекси ДНК з гістонами називають нуклеосомами. В одну нуклеосому входить 8 молекул гістонів і ділянка ДНК, яка містить близько 150 пар основ. Між нуклеосомами є ділянки спіралі, в яких містяться 30—100 пар азотистих основ без суперспіральної структури.
Часто утворюють суперспіраль кільцеві дволанцюгові ДНК, які закручуються самі на себе. Такі суперспіралі характерні для ДНІ< деяких онкогенних вірусів і нехромосомних ДНК цитоплазми бактерій (плазмід) та мітохондрій.
РНК виявлені майже в усіх клітинних фракціях. Найбільша кількість РНК зосереджена в рибонуклеопротеїд- них частинах — рибосомах, де вміст РНК в окремих випадках становить майже 50 %. Взагалі основна маса клітинної РНК міститься в цитоплазмі і лише 10—20 % її виявлено в ядрі.
Рибонуклеїнові кислоти залежно від їх функцій, молекулярної маси і локалізації в клітині поділяють на такі основні групи:
Транспортні РНК (тРНК). Транспортні РНК, або, як їх раніше називали, розчинні РНК (sPHK) від анг. soluble — розчинний) становлять 10—15 % всієї РНК клітини. Вони локалізовані головним чином в гіалоплазмі клітини, ядерному соці і безструктурній частині мітохондрій і хлоропластів.
Характерною ознакою тРНК є їх відносно невелика молекулярна маса.
Основна функція тРНК — кодування і перенесення активованих амінокислот від комплексу амінокислота-АМФ- фермент до місця біосинтезу білка, тобто до рибосом. Зв’язування амінокислот тРНК відбувається за рахунок складноефірного зв’язку. Він утворюється внаслідок взаємодії карбоксильної групи амінокислоти з 3-гідроксилом кінцевого залишку АМФ полінуклеотидного ланцюга тРНК.
Дослідження тРНК показало, що вони мають високу специфічність. Кожна амінокислота має свою тРНК. Кількість різних тРНК в клітині має дорівнювати кількості амінокислот. Останнім часом виявлено, що одна амінокислота може мати дві й більше своїх тРНК. На сьогодні ін- дентифіковано понад 60 різних тРНК.
Рибосомні РНК (рРНК). Рибосомні рибонуклеїнові кислоти зосереджені в рибосомах, де вони міцно зв’язані з білком. Для рРНК характерна висока відносна молекулярна маса. Вона коливається для переважної більшості їх у межах 500 тис.— 2 млн.
Відомо кілька видів рРНК з різними коефіцієнтами седиментації. Наприклад, у рибосомах, що виділені з кишечної палички і складаються з 50S- і 30S-субодиниць, виявлено 23S-PHK з відносною молекулярною масою 1,1 · 106 і 16S-PHK з відносною молекулярною масою 0,6-106.
У складі рибосом клітин тваринного походження також виявлено два види pPHK:18S-PHK і 28S-PHK, які мають різні відносні молекулярні маси (0,7-106 і відповідно 1,8·106).
Поряд з високомолекулярними, у рибосомах виявлено і низькомолекулярні рРНК. Наприклад 5S-PHK, яка містить близько 120 нуклеотидних залишків і 7S-PHK—містить близько 130 нуклеотидних залишків. Всі види рРНК є структурною основою рибосом. Функція різних рРНК вивчена ще недостатньо.
Інформаційна або матрична РНК (іРНК, або мРНК). Інформаційні РНК становлять лише близько 5 % всієї РНК. Кожна молекула іРНК дістає в процесі свого синтезу в ядрі інформацію від ДНК у формі скопійованої послідовності азотистих основ і переносить її на рибосоми, де вона реалізується в процесі біосинтезу білка. Кожний з тисячі білків, які синтезуються в клітині, кодується специфічною іРНК або специфічною ділянкою іРНК.
ІРНК завжди з’єднується іонними зв’язками з білком, утворюючи рибонуклеопротеїдні частинки.
Вірусні Р Н К є складовими частинками вірусних і фагових рибонуклеотидів і несуть всю інформацію, необхідну для розмноження віруса в клітинах хазяїна. їм властива висока відносна молекулярна маса. Деякі віруси мають дволанцюгові РНК.
Для ЕНК, як і для ДНК, характерні кілька рівнів структури.
Первинна структура РНК- Основу хімічної будови РНК становлять полінуклеотиди різної довжини. Порядок чергування залишків відповідних нуклеотидів у ланцюгах характеризує первинну структуру РНК.
Дослідження первинної структури різних видів РНК показало, що для них характерна в основному така сама закономірність у нуклеотидних співвідношеннях, як і для ДНК. При цьому слід враховувати, що до складу РНК з азотистих основ замість тиміну входить урацил.
Крім того, у РНК сума пуринових основ не завжди дорівнює сумі піримідинових основ. їх співвідношення в РНК змінюється в широких межах. Для РНК порівняно з ДНК менш виражена і видова специфічність.
Зараз досягнуто певних успіхів у вивченні первинної структури транспортних, рибосомних та інформаційних РНК.
Вивчення первинної структури тРНК показало, що вони складаються переважно з 70—90 нуклеотидних залишків і мають певні спільні ознаки. Так, на 5'-кінці полінуклеотидного ланцюга у переважній більшості випадків міститься залишок гуанозинмонофосфорної (ГМФ), а на З'-кінці — фрагмент, що складається з двох залишків цитидинмонофосфорної кислоти і одного залишку аденозинмонофосфорної кислоти (ЦМФ, ЦМФ, АМФ). Між ними в полінуклеотидному ланцюгу в точно визначеній послідовності розміщені всі інші пуринові і піримідинові нуклеотид- ні залишки. Серед них близько 8—10% нуклеотидів, які містять мінорні основи: псевдоуридин, різні метильовані похідні аденіну, гуаніну, цитозину та ін. Найбільш поширеними мінорними основами в нуклеотидних тРНК є псевдоуридин і дигідроуридин. Вважають, що мінорні основи підвищують стійкість тРНК проти гідролізу при дії на них лугів і рибонуклеази. Крім того, окремі мінорні основи беруть участь в кодуванні амінокислот, а також мають важливе значення для розпізнавання ферментом аміноацил-тРНК-синтетазою тієї транспортної РНК, яка взаємодіє з певною амінокислотою в процесі її активування.
Вперше встановив первинну структуру тРНК американський дослідник Р. Холлі із співробітниками в 1965 р. Це була тРНК, яка переносить амінокислоту аланін, тобто аланінова тРНК (тРНКала). Значний внесок у вивчення первинної структури тРНК зробили радянські вчені, зокрема О. О. Баєв і його співробітники. Вони повністю розшифрували первинну структуру тирозинової РНК (тРНКтир).
Тепер уже вивчена первинна структура більш як 50 тРНК. Назва цих тРНК походить від амінокислот, які вони переносять до місця біосинтезу білка.
Успішно проводяться дослідження з вивчення первинної структури інших видів РНК. Так, добре вивчена первинна структура різних видів низькомолекулярних рибосомаль- них РНК- Дослідження нуклеотидного складу 5S-pPHK показало, що в цьому виді рибосомальних РНК майже відсутні метильовані азотисті основи і лише в окремих випадках знайдена псевдоуридинова кислота. Співвідношення основних нуклеотидів характеризується явно вираженим ГЦ-типом. В усіх дослідженнях 5S-pPHK, за невеликим винятком, знайдено 120 нуклеотидних залишків.
Значним досягненням є вивчення первинної структури 16S-pPHK, яка містить близько двох тисяч нуклеотидних залишків.
Про первинну структуру мРНК відомо ще досить мало. Вони являють собою одноланцюгові полінуклеотиди, які разом з інформаційними містять окремі неінформаційні ділянки. З неінформаційних ділянок вивчені поліаденілові фрагменти, які містять від 50 до 400 нуклеотидних залишків, і розміщені в кінці молекули мРНК. Нині розшифрована третинна структура мРНК бактеріофага MS2. Ця мРНК містить 3569 нуклеотидних залишків. На початку молекули і в ЇЇ кінці розміщені дві неінформаційні ділянки. Перша з них містить 129 нуклеотидних залишків, а друга—174.
Нині вивчається нуклеотидна послідовність високомо- лекулярних рибосомальних і вірусних РНК. Уже встановлена первинна структура фрагментів РНК фагів R17 Ml6S-pPHK.
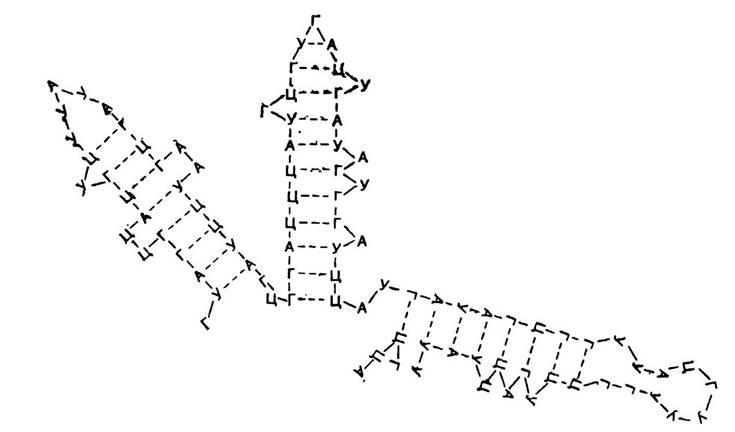
Мал. 12. Вторинна структура РНК.
Вторинна структура РНК. Більшість видів РНК, на відміну від ДНК, складаються з одного полінуклеотидного ланцюга, для якого властива також своєрідна спіралізація. Полінуклеотидний ланцюг РНК закручується сам на себе, утворюючи водневі зв’язки між азотистими основами аденін-урацил і гуанін-цитозин (мал. 12),
Характерною особливістю вторинної структури РНК є те, що полінуклеотидний ланцюг спіралізований не повністю. Крім того, на відміну від ДНК, спіралізація окремих ділянок полінуклеотидного ланцюга РНК менш досконала. У РНК немає повної відповідності в чергуванні комплементарних основ., а тому окремі нуклеотиди виступають на поверхні спіралі полінуклеотидного ланцюга.
Кількість і величина спіралізованих ділянок у межах одного ланцюга для різних РНК неоднакова. Низький ступінь спіралізації властивий для мРНК, що, очевидно, пов’язано з їх функцією в процесі біосинтезу білка. Наявність значної кількості спіралізованих ділянок утруднювало б виконання ними функцій матриці при синтезі поліпептид- ного ланцюга на рибосомах. Водночас на початку полінуклеотидних ланцюгів окремих мРНК виявлено обширні спіральні ділянки.
Вищий ступінь спіралізації (більш як 50 %) властивий для рибосомальних І транспортних РНК. Так, дослідження вторинної структури 5S-pPHK різних видів бактерій показало, що вона містить значну кількість біспіральних ділянок.
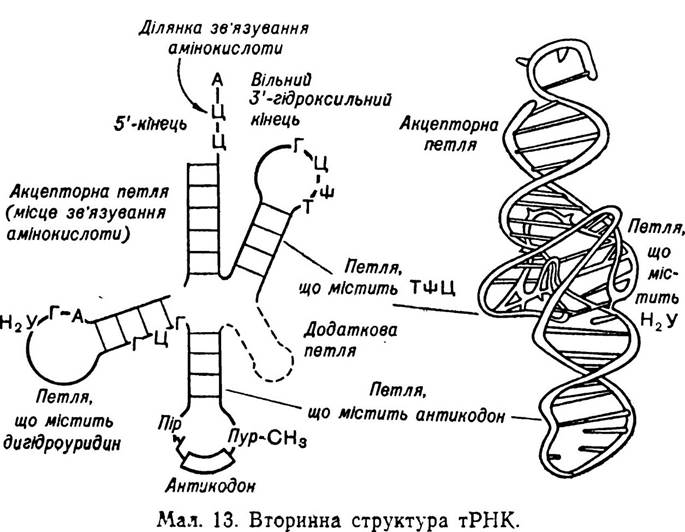
Значно краще вивчена вторинна структура тРНК. Вона нагадує форму листка конюшини (мал. 13), в якому розрізняють такі основні ділянки: 1) центральну ділянку (акцепторну ділянку), яка відповідає за приєднання залишків амінокислот. В усіх тРНК вона однакова і закінчується триплетом — ЦМФ, ЦМФ, АМФ; 2) антикодонову петлю, що містить антикодон — триплет з нуклеотидним чергуванням комплементарного кодону мРНК. Нуклеотиди, які утворюють антикодон, завжди розміщені посередині петлі; 3) псевдоуридинову петлю (Т — φ— Ц-петля), яка завжди містить мінорний нуклеотид— псевдоурйдил (φ), що зв’язаний також мінорною основою для РНК — тиміном. Вважають, що саме ця петля відіграє важливу роль у взаємодії тРНК з рибосомою (50S-субодиницею); 4) дигідроуридилову петлю, яка, очевидно, має певне значення для розпізнавання тРНК своїм ферментом — аміноацилсинтетазою. Звичайно, важливу роль в цьому відіграють і інші ділянки тРНК. Деякі дослідники вважають, що в цьому процесі важливе значення має наявність мінорних основ; 5) додаткову петлю (S-ділянка), біологічна роль якої вивчена ще недостатньо. У деяких тРНК вона відсутня.
Структура листка конюшини, очевидно, характерна для всіх транспортних РНК, в яких уже вивчена первинна структура.
В деяких вірусів (вірус раневих пухлин рослин, реовірус), виявлені дволанцюгові РНК. Дволанцюгові РНК утворюються в процесі розмноження тих вірусів, які містять одно- ланцюгові РНК. їх називають реплікативними дволанцюговими РНК.
Вони виконують роль матриці для вірусних РНК — полімераз. Дволанцюгові РНК виявлені і в евкаріотів.
Проте їх функція і походження ще не вивчені.
За ознаками структурної організації дволанцюгові РНК вірусів нагадують ДНК і є носіями спадкової інформації. Разом з цим вважають, що вторинна структура дволанцюгових РНК має свої особливості. Так, деякі вірусні РНК відрізняються від ДНК кількістю нуклеотидних залишків на один виток, величиною відхилення азотистих основ від перпендикуляра до осі спіралі, відстанню між парами основ тощо.
Особливості структури дволанцюгових РНК, очевидно, зумовлюють їх вищу стійкість порівняно з дволанцюговими ДНК. Наприклад, дволанцюгові РНК, які містять однакову кількість Г — Ц-нар з ДНК, мають вищу температуру плавлення. Для першої вона становить 99 °С, а для другої — 87 °С.
Третинна структура РНК порівняно з ДНК вивчена краще. Так, О. С. Спірін та інші вчені довели, що макромолекули рибосомальної РНК можуть змінювати свій стан і третинну структуру залежно від умов середовища— температури, іонної сили, pH та ін. (мал. 14). При кімнатній температурі і низькій іонній силі рРНК набирає форму палички, а з підвищенням іонної сили — форму компактного клубка. У безсольовому середовищі і при високій іонній силі рРНК набирає форму розгорнутої нитки або рихлого клубка. Третинну структуру, подібну до рибосомальної, за певних умов мають і матричні РНК.
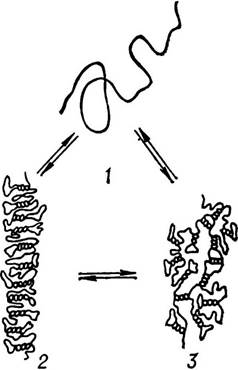
Мал. 14. Третинна структура РНК вірусу тютюнової мозаїки (за О. С. Спіріним та ін.).
t — розгорнута нитка; 2 — компактна паличка; 3 — компактний клубок.
Досить детально вивчена третинна структура деяких видів транспортних РНК. Так, добуті дані свідчать що нативні молекули тРНК мають приблизно однакову третинну структуру. Вона відрізняється від плоскої структури листка конюшини більшою компактністю внаслідок складання різних частин молекули.
Властивості нуклеїнових кислот
Нуклеїнові кислоти — речовини білого кольору, волокнистої будови, погано розчиняються в воді. їх солі з лужними металами добре розчинні у воді. Нуклеїнові кислоти розчиняються в розчинах солей: РНК — у розведених, а ДНК — у більш концентрованих.
Оскільки молекули нуклеїнових кислот значною мірою асиметричні, їх розчини мають високу в’язкість, яка є важливою характеристикою дволанцюгових ДНК, зокрема їх відносної молекулярної маси. Так, відносна в’язкість 0,01-процентного розчину ДНК з відносною молекулярною масою 5-Ю6 близька до 1,5. Приблизно таку ж в’язкість мають розчини ДНК, в яких вміст нуклеїнової кислоти в п’ять разів більший, а відносна молекулярна маса значно менша — 0,3·106. Руйнування водневих зв’язків в ДНК і розпад її на два полінуклеотидні ланцюги призводить до зниження в’язкості розчинів.
Для нуклеїнових кислот характерна також висока оптична активність. їх розчинам властивий значний позитивний поворот площини поляризованого світла. Він помітно зменшується із зменшенням ступеня впорядкованості полінуклеотидних ланцюгів. Так, питома активність розчинів біспіральної ДНК дорівнює 150°. Для розчинів мономер- них нуклеотидів, одноланцюгових РНК і ДНК при однаковій довжині хвилі питома активність в чотири — шість разів менша. І навпаки — при переході від мононуклеотидів до полінуклеотидів питома активність розчину зростає. Це пов’язано із збільшенням асиметрії молекул і ступенем спіралізації полінуклеотидних ланцюгів.
Усі нуклеїнові кислоти мають здатність поглинати світло в ультрафіолетовій області з максимум поглинання 200 нм. Порушення нативності нуклеїнових кислот супроводжується підвищенням поглинання світла, тобто має місце так званий гіпохромний ефект. Він безпосередньо залежить від вмісту в складі нуклеїнових кислот азотистих пар (А+Т). Найбільш помітно гіпохромний ефект виражений для дволанцюгових нуклеїнових кислот, зокрема ДНК. Наявність гіпохромності — одна з важливих ознак утворення дволанцюгових, спіралізоваиих ділянок в нуклеїнових кислотах. Одноланцюгові нуклеїнові кислоти, в яких спіралізація нуклеотидних ланцюгів незначна, мають і незначний гіпохромний ефект. Гіпохромний ефект широко застосовується під час вивчення процесів денатурації і ренатурації нуклеїнових кислот, для утворення гібридних спіралей ДНК—РНК та ін.
Денатурація і ренатурація нуклеїнових кислот. Подібно до білків нуклеїнові кислоти мають здатність до денатурації. Це процес розриву водневих і вандерваальсових зв’язків, деспіралізації та розходження полінуклеотидних ланцюгів ДНК і двоспіральних ділянок молекул РНК. Основними факторами, які зумовлюють денатурацію, може бути дія кислот, лугів, спиртів, нагрівання тощо. Внаслідок денатурації кожний з полінуклеотидних ланцюгів молекули нуклеїнової кислоти набирає форми клубка, згорнутого без належного порядку. Тому такий процес ще називають переходом спіраль-клубок.
Якщо нагріти розчин ДНК до певної температури, можна спостерігати різке підвищення поглинання світла при довжині хвилі 260 нм. Цю температуру називають температурою плавлення ДНК- Вона підвищується із збільшенням довжини полінуклеотиду, а також із підвищенням в ньому вмісту пар азотистих основ Г—Ц. Вважають, що пари азотистих основ в молекулах ДНК утворюють термостабільні ланки, або ядра, і підвищують їх стійкість проти денатурації. І навпаки — молекули ДНК, які містять більшу кількість азотистих основ А—Т, легше піддаються денатурації. Це пояснюється тим, що пара азотистих основ Г—Ц утворює три водневих зв’язки, а азотисті основи А—Т — лише два.
Для вивчення біологічної ролі і функцій ДНК в організмі важливе значення має з’ясування причин і умов, за яких може відбуватися відновлення нативної двоспіральної структури ДНК. Доведено, що при підвищенні температури приблизно на 5 °С вище від температури плавлення ланцюги денатурованої ДНК розходяться. Якщо розчин таких ДНК раптово охолодити, то полінуклеотидні ланцюги так і залишаються розділеними. А якщо охолодження розчину проводити поступово, внаслідок рекомбінації відбувається ренатурація (відновлення) подвійної спіралі ДНК (мал. 15).
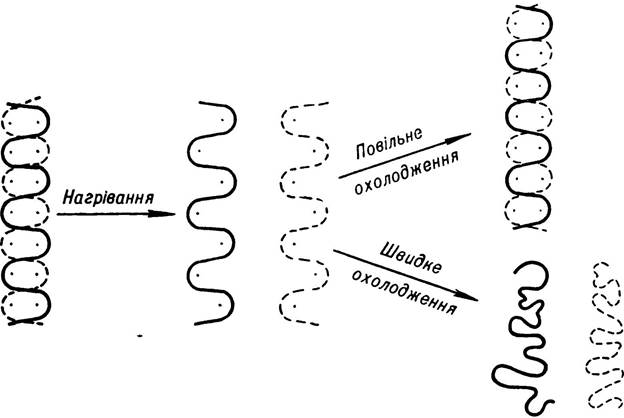
Мал. 15. Денатурація ДНК при нагріванні і вплив охолодження на нагрітий розчин.
Процес ренатурації молекул ДНК залежить від їх розмірів, кількості азотистих пар Г—Ц та інших факторів. Так, ДНК вірусів і бактерій, яка за своєю будовою простіша, ніж ДНК вищих організмів, піддається ренатурації значно легше. Позитивно впливає на процес ренатурації також підвищення кількості Г—Ц-пар.
При ренатурації ДНК відновлюються її біологічні та фізико-хімічні властивості.
Гібридизація ДНК. Встановлено, що полінуклеотидні ланцюги денатурованих ДНК можуть взаємодіяти на основі принципу комплементарності з одноланцюговими РНК і ДНК, які належать іншим організмам. Такий вид ренатурації називається молекулярною гібридизацією. Вважають, що для утворення гібридної молекули ДНК достатньо в полінуклеотидних ланцюгах до 12 комплементарних пар азотистих основ. Чим більше комплементарних пар, тим більший вміст біспіральних ділянок в гібридній молекулі ДНК.
Відкриття процесу гібридизації ДНК має важливе значення для вивчення первинної структури різних видів нуклеїнових кислот, а також сприяє науковим дослідженням в галузі генної інженерії.
Хімічні реакції нуклеїнових кислот. Мутагени. Оскільки в складі основ і пентоз нуклеїнових кислот є різні функціональні групи, вони можуть вступати в хімічні реакції, що призводить до видозмінення основ і впливає на структуру і функції нуклеїнових кислот.
Так, аміногрупи азотистих основ можуть взаємодіяти з азотистою кислотою. При цьому замість групи — NH2 утворюється група — ОН.
Азотиста кислота, взаємодіючи з нуклеїновими кислотами, перетворює цитозин в урацил, а аденін і гуанін відповідно в гіпоксантин і ксантин. Отже, вона є досить ефективним хімічним мутагеном.
Аналогічну мутагенну дію має гідроксиламін, який вступає в реакцію з карбонільними групами азотистих основ, зокрема з піримідиновими, що приводить до перетворення одного виду основ в інші.
Високу мутагенну дію мають алкілуючі агенти (нітрозаміни). Вони сильні канцерогени. Нітрозаміни можуть утворюватися від взаємодії будь-якого вторинного аміну з азотистою кислотою:
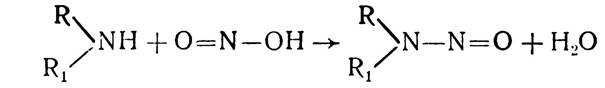
Така реакція може відбуватися і в шлунку людини, де нітрозаміни легко всмоктуються його стінками і можуть спричинити розвиток злоякісних пухлин у різних органах і тканинах організму. Нуклеїнові кислоти можуть взаємодіяти з окремими поліциклічними ароматичними (основами) барвниками, зокрема з акридиновим оранжевим, профлавіном та ін., катіони яких мають плоску будову. Вибірково зв’язуючись з ДНК або РНК, вони дають можливість легко їх виявляти. Останнім часом ці барвники набули важливого значення як селективні ' мутагени. Вони здатні викликати вставки або делеції (включення або випадання окремих одиночних нуклеотидів) при реплікації ДНК і, таким чином, пригнічуючи її.
Нуклеїнові кислоти можуть гідролізуватися під впливом кислот. Глікозидні зв’язки в ДНК більш лабільні, ніж в РНК. Нуклеотиди, які містять пуринові основи, гідролізуються легше, ніж нуклеотиди з піримідиновими основами. І все-таки, незважаючи на здатність до гідролізу, ковалентна структура молекул нуклеїнових кислот досить стійка, що дає можливість їм виконувати функцію генетичного матеріалу.
Репарація пошкоджень ДНК. Пошкодження ДНК, які зумовлені дією різних хімічних і фізичних факторів, можуть бути репаровані (виправлені) з участю спеціальних механізмів «ремонтних систем». Наприклад, при дії ультрафіолетового опромінювання на ДНК утворюються тимінові димери внаслідок взаємодії в одному ланцюгу ДНК сусідніх тиміновнх залишків з утворенням циклобутанового кільця.
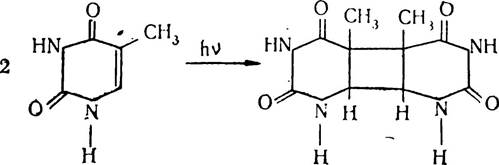
Утворення тиміновнх димерів порушує структуру ДНК і тим самим блокує її реплікацію. Цим, очевидно, можна пояснити летальну та мутагенну дію ультрафіолетового .опромінення на живі організми.
Для виправлення пошкоджень ДНК в живих системах існує кілька механізмів, зокрема фотореактивація, ексцизійна і постреплікативна репарація.
Фотореактивація відбувається внаслідок дії видимого світла. При цьому відбувається активація фотореактивного фермента (ДНК-фотолази), який розщеплює димери на мономери і відновлює водневі зв’язки між тиміном і аденіном комплементарних полінуклеотидних ланцюгів ДНК.
Ексцизійна і постреплікативна репарація не залежить від світла і тому її ще називають темновою репарацією. Ці два види репарації подібні між собою.
При ексцизійній репарації (лат. excisio — вирізання) відбувається видалення пошкодженої ділянки ДНК з участю цілого комплексу ферментних систем. Так, спочатку діє фермент ендонуклеаза, що розрізає полінуклеотидний ланцюг ДНК, на якому є пошкодження нуклеотидів. Потім вступає в дію фермент екзонуклеаза, яка видаляє пошкоджену частину нуклеотидного ланцюга. На цьому місці з участю ферменту ДНК-полімерази в напрямі 5->3 заново синтезується така сама частина комплементарного полінуклеотидного ланцюга з непошкодженими нуклеотидами. На заключному етапі вільні кінці старої частини полінуклеотидного ланцюга з участю ферменту лігази з’єднуються з кінцями синтезованої частини полінуклеотидного ланцюга.
Отже, клітини організмів містять цілі набори ферментів, які переміщаються по подвійних спіралях ДНК, відновлюють їх пошкодження і цим самим сприяють зменшенню появи частоти мутацій.
Матричний синтез нуклеїнових кислот.
Біосинтез ДНК. Процес біосинтезу ДНК ще називають реплікацією. Реплікація ДНК відбувається напередодні кожного нормального поділу ДНК-вмісних структур (ядер, мітохондрій, пластид) у евкаріотів і перед кожним поділом бактеріальних клітин та при розмноженні ДНК-вірусів.
Вихідними сполуками для синтезу ДНК є дезоксирибонуклеотид-5'-трифосфати. Сумарну реакцію синтезу ДНК можна показати у вигляді такої схеми:
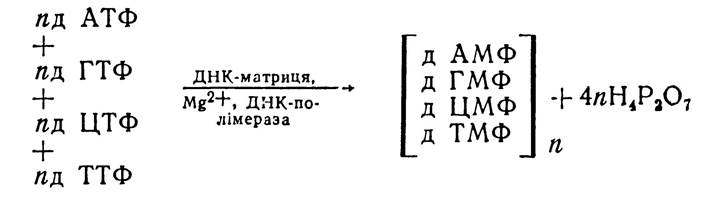
У вигляді матриці для синтезу ДНК використовується дволанцюгова ДНК клітин. До обох її полінуклеотидних ланцюгів добудовуються нові полінуклеотидні ланцюги на основі принципу комплементарності. Реплікація відбувається напівконсервативним способом. Отже, в результаті реплікації з однієї молекули дволанцюгової ДНК утворюються дві однакові молекули ДНК, які також ідентичні з вихідною молекулою ДНК. Це є передумовою збереження видоспецифічної спадкової інформації в ряді поколінь клітин та організмів, а також забезпечення постійності видів.
Весь процес біосинтезу ДНК поділяють на такі етапи: ініціацію, елонгацію і термінацію.
Ініціація полягає в розкручуванні двоспіральної структури ДНК і поступовому розходженні полінуклеотидних ланцюгів з утворенням так званої реплікативної вилки. Цей процес відбувається з участю ДНК-зв’язуючого і ДНК-розкручуючого білків та ДНК-полімеразного комплексу. Важливе значення для початку синтезу ДНК має наявність ферменту РНК-полімерази або примази. З його участю відбувається синтез невеликої ділянки (до 50 нуклеотидних залишків) РНК, яка є затравкою (праймером) синтезу ДНК. Вона може утворюватись в кількох ділянках полінуклеотидних ланцюгів реплікативної вилки. Це зумовлює той факт, що реплікація відбувається не безперервно, а окремими фрагментами, які називаються фрагментами Оказаки на честь японського дослідника.
Елонгація полягає в реплікації ділянок материнських ланцюгів ДНК і з’єднанні новоутворених ділянок (фрагментів Оказаки) між собою.
Реплікація полінуклеотидного ланцюга ДНК починається з приєднання до 3"-гідрокеилу затравки дезоксинуклеозидфосфату, до якого приєднується нова молекула дез- оксинуклеозидфосфату і так продовжується доти, доки новий полінуклеотидний ланцюг не дійде до місця, де розміщена нова затравка. З неї починається ріст наступного (нового) фрагмента полінуклеотидного ланцюга. Такі фрагменти можуть складатися з 1000—2000 нуклеотидних залишків. Процес реплікації відбувається з участю ферменту ДНК-полімерази III, яка каталізує ріст полінуклеотидного ланцюга в напрямі 5→3.
За даними Оказаки, у живих клітинах має місце човниковий механізм реплікації. Суть його полягає в тому, що спочатку реплікація відбувається в напрямі 5 →3 просуванням вперед по одному ланцюгу, а потім в тому самому напрямі пересувається в зворотний бік по іншому полі- нуклеотидному ланцюгу материнської ДНК, тобто один полінуклеотидний ланцюг синтезується в напрямі репліка- ційної вилки, а інший — від неї.
Синтезовані фрагменти Оказаки на 5"-кінці дезоксирибози містять ковалентно зв’язану РНК-затравку. Остання з участю ДНК-полімерази І, яка має нуклеазну активність, видаляється, і її місце заповнюється синтезованим ланцюгом ДНК. Потім фрагменти Оказаки при каталітичній дії ферменту лігази зв’язуються між собою.
Вважають, що реплікації піддаються одночасно обидва полінуклеотидні ланцюги ДНК. При цьому один з ланцюгів, які синтезуються, є ведучим, а інший — запізнюючнм.
Термінація (останній етап синтезу ДНК) відбувається внаслідок закінчення росту полінуклеотидних залишків ДНК матриці. Цей процес відбувається за рахунок нуклеотидилтрансферазної реакції. Реплікація ДНК в евкаріотів складніша, ніж у прокаріотів. Швидкість реплікації у прокаріотів становить 1000—2000 нуклеотидів за секунду і близько 100 нуклеотидів за цей час в евкаріотів.
Процес реплікації відбувається з великою точністю. Так, на 10'° пар нуклеотидів зустрічається лише одна помилка. Включення неправильного нуклеотиду, наприклад урацилу замість тиміну, відразу ж зумовлює його видалення із синтезованого полінуклеотидного ланцюга і введення на його місце тиміну.
Забезпечення точності реплікації — один з важливих етапів передачі спадкової інформації від одного покоління до іншого.
РНК — залежний біосинтез ДНК. Протягом останнього десятиріччя розроблено спосіб синтезу ДНК та РНК з участю ферменту, який названо зворотною тран- скриптазою, або ревертазою. Цей фермент ще називають РНК-залежною-ДНК-полімеразою. Він був виділений у 1969—1970 роках із тканин, інфікованих онкогенними вірусами, а також із самих вірусів цієї групи, зокрема з вірусу Раушера лейкемії мишей і вірусу саркоми Рауса. Реверта- за активується катіонами марганцю (II) і магнію, причому катіонами марганцю (II) ефективніше.
Вивчення будови ферменту, виділеного з вірусів мієлобластоза птахів, показало, що він складається з двох білкових субодиниць з відносною молекулярною масою 110 і 70 тис. і містить два атоми зв’язаного цинку. Для функціонування ферменту ревертази необхідні затравка і матричний ланцюг РНК. Спочатку на одноланцюговій РНК-матриці з участю ферменту ревертази синтезується одноланцюгова ДНК. Внаслідок цього утворюється гібридна біспіральна РНК-ДНК молекула, яка потім виконує роль матриці для синтезу комплементарного полінуклеотидного ланцюга ДНК. Цей етап каталізується ферментом РНК-залежною-ДНК-полімеразою. На останньому етапі з участю ферменту РНК-ази відбувається відщеплення молекули ДНК від РНК. Потім молекула ДНК включається в геном інфікованої клітини і спричиняє переродження її в ракову.
Наявність зворотної транскриптази встановлено в культурах синцитіальних вірусів, які не зумовлюють злоякісного переродження клітин. У певних умовах здатність каталізувати синтез ДНК на РНК мають і ДНК-полімераза І та евкаріотичні ДНК-полімерази.
Відкриття процесу зворотної транскрипції має важливе теоретичне і практичне значення. Так, якщо раніше вважалось, що передача генетичної інформації в усіх організмів, за винятком РНК-вмісних вірусів, відбувається лише від ДНК до ДНК і від ДНК до РНК, то зараз встановлено, що вона може здійснюватись і в напрямі від РНК до ДНК. Виявлення ферменту ревертази має важливе значення для ранньої діагностики ракових захворювань та в генній інженерії для синтезу окремих геній.
Біосинтез РНК на матриці Д Н К. На матриці дволанцюгової ДНК синтезується переважна більшість РНК людини, тварин, бактерій і ДНК-вмісних вірусів. Цей процес відбувається також за принципом комплементарності азотистих основ нуклеотидів. Відмінність полягає лише в тому, що замість азотистої основи тиміну, яка характерна для ДНК, в РНК включається урацил. Для синтезу РНК на ДНК необхідні всі чотири види нуклеотидів у формі трифосфатів. Процес синтезу можна показати у вигляді такої схеми:
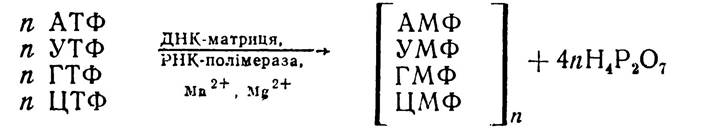
Реакцію каталізують ферменти РНК-полімерази. Тепер добре вивчений фермент типу РНК-полімераза кишкової палички. Відносна молекулярна маса її становить 480 тис. Фермент складається з чотирьох субодиниць: α, β, β', γ, які мають різну відносну молекулярну масу. Вони входять до складу РНК-полімерази в кількісному співвідношенні 2:1:1:1. Комплекс, який містить α, β, β'-субодиниці, називається корферментом. Для утворення активної форми ферменту необхідно всі чотири субодиниці.
РНК-полімерази виділені також з культури клітин людини, тимусу телят, печінки щурів. У кожному випадку в ядерній фракції було виділено три різні ферменти: РНК- полімераза 1(A), РНК-полімераза 11(B) і РНК-полімераза III (С), які відрізняються між собою кількістю субодиниць в молекулі і відносною молекулярною масою. Вважають, що РНК-полімераза І каталізує синтез рибосомальних РНК, РНК-полімерази (II)—синтез матричних РНК і РНК- полімераза III, очевидно, відповідальна за синтез транспортних РНК і 5S-PHK.
РНК-полімерази мають два активних центри. Один з них використовується для зв’язування 3"-ОН-групи рибози кінцевого нуклеотиду ланцюга РНК, який синтезується і називається Т-центром. Другий активний центр зв’язує нук- леотиди, які приєднуються до ланцюга РНК, що росте, і називається П-центром.
Процес матричного синтезу РНК з участю РНК-полімераз поділяють на такі стадії: приєднання ферменту до матриці, ініціація синтезу, елонгація — ріст нуклеотидного ланцюга і термінація — завершення синтезу РНК та звільнення ферменту. В цілому всі ці стадії становлять цикл транскрипції.
Приєднання ферменту до матриці. На цій стадії РНК-полімераза взаємодіє з невеликою ділянкою ДНК, утворюючи комплекс, який може починати зв’язування рибонуклеозидтрифосфатів і ініціювати синтез РНК. Хоч РНК-полімераза виявляє велику спорідненість до всіх ділянок ДНК, проте вона найміцніше зв’язується лише ділянками, які характеризуються певними послідовностями нуклеотидів на одному з ланцюгів матричної ДНК. Ці ділянки називають промоторами. Вони містять багато АТ-пар нуклеотидів і піримідинових блоків. Вважають, що специфічність взаємодії РНК-полімерази з певними ділянками ДНК-матриці зумовлена наявністю в складі цих ферментів 6-субодиниць (6-факторів).
Зв’язування РНК-полімерази з промоторною ділянкою ДНК призводить до часткового плавлення (розходження) спіралі ДНК, що важливо для початку транскрипції РНК.
Ініціація. На цій стадії РНК-полімераза з участю Т-центра взаємодіє з 3'-ОН-групою рибози першого нуклео- зидтрифосфату, з якого починається синтез нової рибонуклеїнової кислоти. При цьому від нього не відщеплюється залишок пірофосфату. Першим нуклеозидтрифосфатом в основному є АТФ або ГТФ. Потім РНК-рибонуклеаза другим активним центром (П-центром) взаємодіє з другим нуклеозидтрифосфатом, який з участю фосфодіефірного зв’язку приєднується до 3"-вуглецю рибози першого нуклеозидтрифосфату. При цьому вивільняється Т-центр ферменту і відщеплюється пірофосфат за рахунок другого нуклеозидтрифосфату.
Елонгація. На стадії елонгації перший активний центр ферменту зв’язується з 3"-ОН-групою рибози другого нуклеотиду і РНК-полімераза переміщується (транслокується) на поверхні ДНК в бік зростання ланцюга. За цих умов другий активний центр ферменту зв’язується з наступним (третім) нуклеотидом, який знову з участю фосфодіефірного зв’язку з’єднується з другим нуклеозидмонофосфатом. Так відбувається ріст, видовження (синтез) нуклеотидного ланцюга в напрямі 5→3 до повного закінчення біосинтезу РНК.
Термінація — процес обриву ланцюга порівняно з іншими етапами синтезу РНК менш вивчений. Вважають, що ДНК має специфічні термінуючі послідовності нуклеотидів. РНК-полімераза, доходячи до них, припиняє свою функціональність. При цьому синтезований полінуклеотидний ланцюг РНК і молекула ферменту відділяються від ДНК-матриці.
Усі види РНК (за винятком вірусних) синтезуються на молекулах ДНК. За цих умов спочатку утворюється попередник відповідної РНК з довгим полінуклеотидним ланцюгом. Його називають первинним транскриптом, який потім перетворюється в коротшу РНК. Це перетворення називається процесінгом, або дозріванням РНК.
Воно охоплює ряд етапів: а) ланцюг РНК укорочується внаслідок відщеплення нуклеотидів з його кінців або видалення певної ділянки із середини полінуклеотидного ланцюга з наступним з’єднанням утворених частин. Цей процес називають сплайсінгом; б) до укороченого полінуклеотидного ланцюга приєднуються нові кінцеві послідовності нуклеотидів; в) відбувається модифікація нуклеотидів, наприклад їх гідрування або метилювання.
Первинний транскрипт і проміжні продукти процесінгу відомі під назвою пре-РНК.
Різні види РНК (пре-РНК та інші, ще мало вивчені РНК), які є в ядрі, становлять так звану фракцію гетерогенної ядерної РНК (гя РНК).
Необхідно зазначити, що синтез РНК істотно відрізняється від реплікації ДНК. Насамперед він є консервативним, а не напівконсервативним — продукт синтезу не включає будь-яких компонентів матриці. Цікавим є і той факт, що ферменти синтезу РНК, тобто РНК-полімерази, мають високу специфічність відносно різних матриць і навіть їх окремих ділянок. Крім того, для матричного синтезу РНК непотрібна затравка. Процес синтезу починається з реакції між двома нуклеотидами. І, нарешті, процес транскрипції, на відміну від реплікації, відбувається переважно з одного полінуклеотидного ланцюга ДНК-матриці.
Функції нуклеїнових кислот
Нуклеїнові кислоти в організмі відіграють різноманітні функції, але найважливішими є участь їх у передачі спадкових ознак і процесах біосинтезу білка.
Основними носіями генетичної інформації в більшості організмів є ДНК. Виняток становлять окремі фаги, дрібні віруси тварин і більшість рослинних вірусів, в яких генетичну інформацію несуть молекули РНК.
Основна кількість ДНК клітин зосереджена в ядрах. Ядра тваринних клітин містять близько 2 мг ДНК на 1 г сирої маси тканини. Вміст ДНК в клітинах залежить в основному від кількості хромосомних наборів у них. Бактеріальні клітини містять в сотні разів менше ДНК, ніж тваринні.
Передача генетичної інформації з покоління в покоління відбувається самовідтворенням точних копій батьківських ДНК. Цей процес називається реплікацією, або розмноженням ДНК. Так, під час поділу клітин обидва полінуклеотидні ланцюги ДНК, які спірально обвиті навколо однієї спільної осі, розкручуються і розходяться. У міру того, як кожний з ланцюгів розкручується, він добудовує собі подібний полінуклеотидний ланцюг на основі принципу комплементарності (мал. 16). Механізм реплікації ДНК описано на с. 124—129.
В результаті реплікації з однієї молекули утворюються дві нові цілком однакові молекули ДНК, одна з яких залишається в материнській клітині, а друга переходить у дочірню. Якщо додати, що в ДНК зосереджена спадкова інформація у вигляді триплетного коду, то стає зрозумілою та важлива функція, яку відіграє ДНК в передачі спадкових ознак від покоління до покоління.
Реплікація, як і будь-який процес побудови та розпаду в організмі, відбувається за допомогою цілого ряду ферментів і тісно пов’язана з обміном речовин. Генетична роль ДНК встановлена також проведенням дослідів на бактеріях різних типів з різними властивостями. Наприклад, з одного типу бактерій виділяли чисту ДНК і діяли нею на інший тип бактерій, при цьому бактерії останнього типу набували властивостей, що характерні для типу бактерій, з якого була виділена ДНК.
Отже, ДНК — матеріальний субстрат, за допомогою якого відбувається передача спадкових ознак. Спадкова інформація організму закладена і зберігається в самій молекулярній структурі ДНК, а реалізується (виявляється) в процесі біосинтезу білка.
У біосинтезі білка велику роль відіграють також і різні види РНК. Встановлено, що РНК міститься в усіх клітинах організму. Однак їх кількість залежить від типу клітини, виду тканини, віку і фізіологічного стану організму. У клітинах, які інтенсивно розмножуються і ростуть, вміст РНК високий. У міру старіння організму і зниження темпів росту концентрація РНК в клітинах зменшується. Оскільки процеси росту і розмноження передусім пов’язані із збільшенням білкової маси цитоплазми, то це в достатній мірі свідчить про тісний взаємозв’язок між РНК і біосинтезом білка в клітинах. В результаті численних досліджень з’ясовано, що клітини, в яких відбувається інтенсивний синтез білка, характеризуються особливо високим вмістом РНК. Наприклад, велику кількість РНК знайдено в залозах, які продукують білки — гормони або ферменти (підшлункова, травні залози).
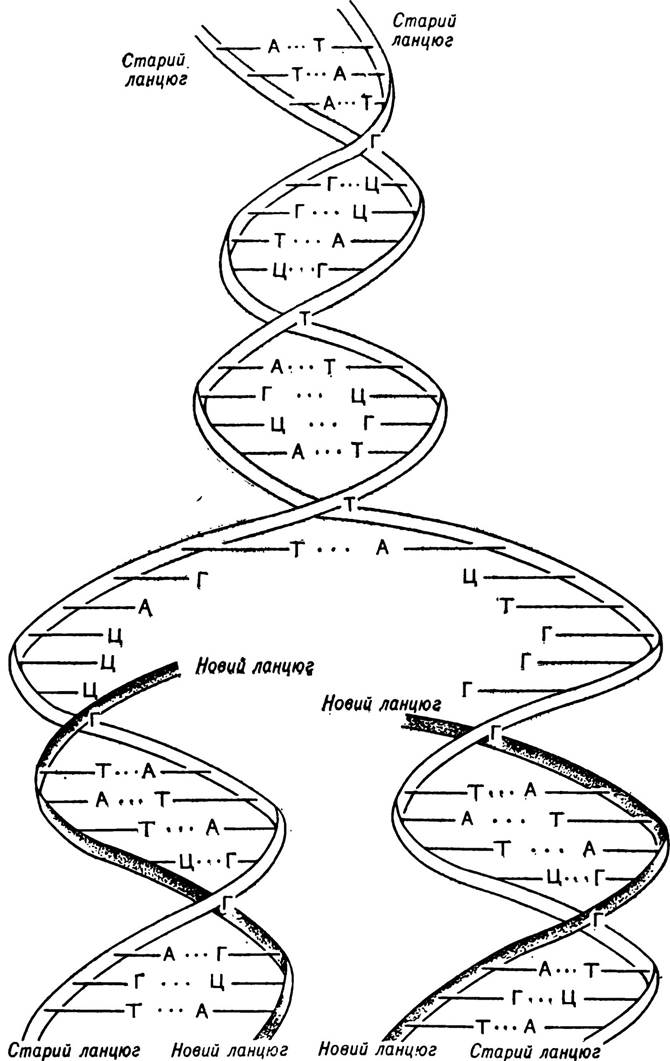
Мал. 16. Схема реплікації ДНК
Слід зазначити, що основна кількість клітинної РНК (до 80 %) зосереджена в рибосомах, тобто в тих компонентах клітин, на яких відбувається біосинтез білка. У рибосомах міститься переважно рибосомальна РНК. Інші види РНК зосереджені частково в ядрі, а частково в цитоплазмі.
Встановлено, що всі три види нуклеїнових кислот беруть участь у біосинтезі білка. При цьому кожний з них виконує свою специфічну функцію. Так, іРНК дістає інформацію про специфічність біосинтезу білка в ядрі від ДНК і переносить її до рибосом, тРНК переносить активовані амінокислоти до місця біосинтезу білка. Роль рибосомних РНК у синтезі ще остаточно не з’ясована. Вважають, що вони створюють клітинну структуру — рибосоми, на яких відбувається синтез білка. Можливо рибосомальні РНК виконують і інші функції у процесі синтезу білка.
ДНК, на відміну від РНК, бере участь у цьому процесі не безпосередньо, а через іРНК. Вона відіграє основну роль у визначенні специфічності біосинтезу білка.