БІОХІМІЯ ДЛЯ ВЧИТЕЛЯ - Ф.Ф. БОЄЧКО - 1985
ОСНОВНІ ХІМІЧНІ КОМПОНЕНТИ КЛІТИН
ГОРМОНИ — РЕГУЛЯТОРИ МЕТАБОЛІЧНИХ ПРОЦЕСІВ
Для забезпечення існування організму як єдиної складної впорядкованої системи з його здатністю до самовідтворення, росту і розвитку необхідним є високий рівень інтеграції і координації всіх життєвих функцій. Щоб існувати, організм повинен швидко реагувати на зміну умов навколишнього середовища, постійно підтримувати динамічну рівновагу всіх метаболічних процесів. Тому в процесі еволюції поступово сформувався централізований апарат управління, основною функцією якого є міжклітинна регуляція і програмування роботи різних органів і тканин організму. Роль такого апарату в більшості тваринних організмів і людини виконують нервова і ендокринна системи.
Ендокринна система виникла на ранніх етапах розвитку тваринного світу, після диференціації нервової системи.
В організмі вищих тварин і людини існує два види залоз — екзокринні (від грецького ekso — назовні, krinein — виділення) та ендокринні (від грецького endo — всередину). Залози зовнішньої секреції (слинні, потові, шлунково-кишкового тракту) виділяють свої секрети безпосередньо в кров.
У хребетних тварин виявлено близько 10 ендокринних залоз, які продукують біологічно активні секрети і становлять ендокринну систему. Найважливіші серед них такі: гіпоталамус, гіпофіз (нижній мозковий придаток), шишкоподібна залоза (епіфіз), щитовидна, тимус, підшлункова, надниркові залози і статеві. Крім того, вважають, що деякі органи, зокрема нирки, печінка і селезінка, містять в своєму складі клітинні елементи, які можна віднести до ендокринних залоз. Ці органи виділяють секрети, які виконують важливу роль у регуляції тонусу судин і ряду обмінних процесів.
Шлунково-кишковий тракт також продукує речовини, які певним чином впливають на окремі процеси метаболізму. Крім цих залоз, які є в більшості класів хребетних, слід згадати особливу ендокринну залозу вагітності ссавців — плаценту, яка продукує велику кількість речовин, що впливають на статеві функції і сприяють протіканню вагітності.
Ендокринних залоз немає в більшості нижчих безхребетних і є лише в найбільш високоорганізованих їх представників— головоногих молюсків, ракоподібних, комах. З участю цих залоз контролюються найважливіші ланки онтогенезу безхребетних — ріст, линяння, метаморфоз, розмноження і адаптація. Секрети, що виділяються залозами внутрішньої секреції, називаються гормонами (від грецького hormanino — приводити в дію, збуджувати). Назву цій групі речовин дали в 1902 р. англійські вчені В. Бейліс і Е. Старлінг на основі дослідження властивостей секретину — речовини, яка посилює секрецію панкреатичного соку. Пізніше з’ясувалось, що гормони є специфічними продуктами ендокринних залоз, які виявляють дистантну регулюючу дію.
Діапазон дії гормонів на метаболічні процеси досить різноманітний — від транспорту іонів через плазматичні мембрани до транскрипції геному. Гормони мають здатність змінювати структури і функції клітин, тканин і органів, їх фізіологію і морфологію. Незаперечним є вплив гормонів на синтез макроергів, ДНК, білків, формування клітинного фенотипу, ріст і розвиток організму, формування статі, адаптацію і підтримання гомеостазу.
Гормони щитовидної залози
Щитовидна залоза — одна з найдосконаліше вивчених залоз внутрішньої секреції. Розміщена вона в області гортані і трахеї і складається з двох лопатеподібних часток, сполучених тонким перешийком. Довжина часток 5—8 см, а ширина 3—4 см. Маса залози 25—ЗО г, це одна з найбільших ендокринних залоз. Тканина щитовидної залози складається з великої кількості залозистих утворень (фолікул), заповнених в’язкою рідиною жовтого кольору (колоїдом), що продукується клітинами залози. Кожний фолікул обплетений густою капілярною сіткою, що забезпечує досить інтенсивний кровообіг у тканинах залози. Маса крові, що проходить через залозу за 1 хв, в 3—5 разів перевищує масу всієї залози. А за 1 год майже вся кров, яка циркулює по судинній системі людини, встигає пройти через цей ендокринний орган.
До складу колоїду, що міститься в фолікулах щитовидної залози, входить складний білок глікопротеїдної природи— тиреоглобулін, що містить 0,5—1 % йоду і 8—10% вуглеводів. Він є необхідною сполукою, з якої утворюються гормони щитовидної залози:
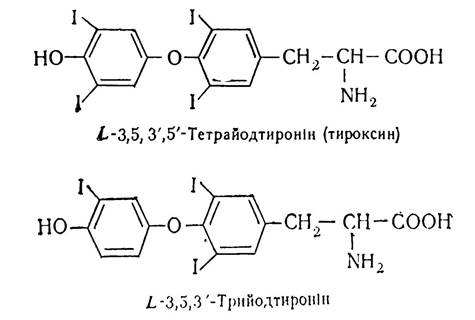
Найбільша біологічна активність властива трийодтироніну, та оскільки його утворюється невелика кількість, специфічна біологічна дія гормону виявляється в основному за рахунок тироксину.
Гормони щитовидної залози є активними регуляторами метаболічних процесів в організмі, вони забезпечують нормальне функціонування багатьох органів і систем організму. Тому при порушенні функцій щитовидної залози в організмі виникає ряд патологічних змін, характер і напрям яких залежить від багатьох факторів.
Основними видами патології при порушенні функціонування щитовидної залози є базедова хвороба, мікседема, ендемічний зоб і кретинізм.
Базедова хвороба. Причиною виникнення цього захворювання є гіперфункція щитовидної залози — надмірне виділення гормонів, що в ній синтезуються. Оскільки багато гормонів, зокрема тироксин, отруйно діють на організм людини, це захворювання називають ще тиреотоксикозом.
Вперше гіперфункцію щитовидної залози описав німецький лікар К. Базедов у 1840 р. Характерною ознакою захворювання є підвищення основного обміну в організмі і посилення процесів розпаду тканинних білків, вуглеводів, жирів внаслідок підвищення активності ферментів, які контролюють ці процеси (гексокіназа, сукцинатдегідрогеназа, тіолові і протеолітичні ферменти). Посилений розпад різних сполук супроводжується нагромадженням великої кількості метаболітів проміжного обміну, для окислення яких використовується велика кількість кисню. Внаслідок цього виникає кисневе голодування, яке негативно впливає на функціонування ряду систем організму, особливо центральної нервової системи.
Тиреотоксикоз — одне з найпоширеніших ендокринних захворювань, яке виникає переважно в жінок 20—40 років. Часто тиреотоксикоз зумовлюється різними психічними травмами. За висловленням С. П. Боткіна, горе, гнів, страх можуть бути причиною важкого тиреотоксикозу, який виникає іноді дуже швидко — протягом кількох годин. Причиною цього є сильне збудження кори головного мозку, яке передається до щитовидної залози безпосередньо по нервових шляхах або через гіпофіз, що зумовлює звільнення великої кількості гормону.
Характерним для тиреотоксикозу є зовнішній вигляд хворих, зокрема, екзофтальм (витрішкуватість), який надає обличчю сердитого і дещо зляканого вигляду (вираз застиглого страху). Хворі різко худіють, за кілька місяців можуть втратити 12—16 кг маси тіла.
Наприкінці минулого століття, коли не було надійних методів лікування цього захворювання, більшість хворих на важку форму тиреотоксикозу помирала протягом 2—З років. Тепер широко використовують препарати, що пригнічують діяльність щитовидної залози (радіоактивний йод, похідні тіоурацилу та імідазолу), а також хірургічні методи.
Мікседема виникає при гіпофункції щитовидної залози, і є діаметрально протилежною базедовій хворобі. Зокрема, гальмуються основні процеси обміну речовин, знижується тиск крові і температура тіла. Хворі стають кволими та апатичними. Спостерігається загальна слабкість, в'ялість, сонливість. Вони постійно мерзнуть, шкіра суха, злущується, особливо на ліктевих згинах і гомілках. Погіршується пам’ять. Температура тіла знижується до 35—36°С, причому, навіть інфекційні захворювання не зумовлюють значного підвищення температури. Спостерігаються слизові набрякання, одутлість обличчя.
Якщо гіпофункція щитовидної залози виникає в дитячому віці або в період внутріутробного розвитку і супроводжується недостачею йоду, виникає захворювання, що' називається кретинізмом. У таких дітей значно затримуються ріст і розвиток. Ходити діти починають пізно — в 2—3 роки, пізно прорізуються зуби. Обличчя в дітей одутле, вираз його байдужий, тупий, рот напіввідкритий, голос грубий, охриплий. Такі діти часто народжуються глухонімими.
При важкому протіканні хвороби з усіх емоцій залишаються лише почуття голоду і болю. Виникає порушення психічної діяльності.
Крім цих захворювань, досить часто виникає ще один вид патології, що називається ендемічним зобом. Причиною виникнення цього захворювання є знижена здатність організму хворого засвоювати йод або недостача його в продуктах харчування і воді. Особливо часто це захворювання зустрічається в гірських та заболочених районах.
Гормони підшлункової залози
Підшлункова залоза — це непарний орган змішаної секреції, який розміщений у черевній порожнині з лівого боку шлунка. Структура цієї залози досить складна. Екзогенну функцію підшлункової залози виконує адипозна (зимогенна) тканина, яка продукує безпосередньо в дванадцятипалу кишку панкреатичний сік.
Внутрісекреторна (ендогенна) функція забезпечується острівками Лангерганса, що являють собою скупчення клітин різного типу — α,β, D. Загальна маса острівкових клітин у людини становить 1/30—1/100 частину маси всієї підшлункової залози.
Острівкова тканина підшлункової залози характеризується постійністю клітинного складу і містить 80 % α-клітин, 20 % β-клітин і 2—8 % D-клітин. Кожний вид клітин продукує певні характерні для нього гормони, які регулюють відповідні ланки метаболічних процесів в організмі людини і тварин.
β-Клітини містять іони цинку і продукують гормон інсулін (insyla — острів), що є важливим регулятором вуглеводного, ліпідного і білкового обміну. В 1 кг підшлункової залози міститься близько 150 мг інсуліну. β-Клітини підшлункової залози можуть вибірково пошкоджуватись алоксаном, солями магнію, дегідроаскорбіновою кислотою.
α-Клітини також містять іони цинку і продукують гормон глюкагон (гіперглікеміко-глікогенолітичний фактор, ГГФ). Обидва гармони з цинком утворюють хелатні комплекси. У D-клітинах (дефінітивних) утворюються, очевидно, три гормони — соматостатин, панкреагастрин і секретин. У підшлунковій залозі містяться також гормоноподібні речовини — ліпокаїн, центропненін, ваготонін.
Інсулін — гормон, що продукується p-клітинами підшлункової залози. Його було відкрито як фактор, що зменшує гіперглікемію у тварин з видаленою підшлунковою залозою.
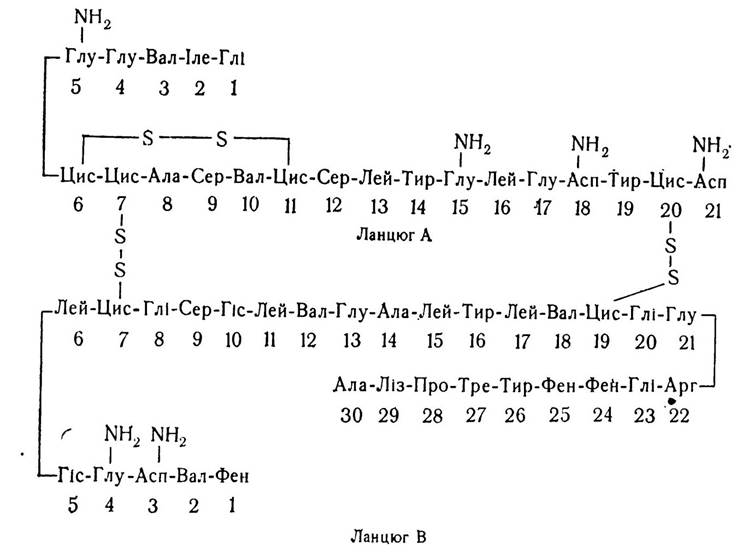
За хімічною природою інсулін є глобулярним білком з відносною молекулярною масою 6000. Молекула інсуліну складається з двох поліпептидних ланцюгів А і В, які містять відповідно 21 і 30 амінокислотних залишків, з’єднаних двома дисульфідними містками, що утворюються між залишками цистеїну.
Синтезується інсулін у^рибосомальному апараті p-клітин острівків Лангерганса у вигляді неактивного попередника (проінсуліну) з відносною молекулярною масою 10 000. Цей попередник містить 78—84 амінокислотних залишків, залежно від виду тварин. У секреторних гранулах під впливом протеолітичних ферментів проінсулін розщеплюється на С-пептид і активну форму інсуліну. Інсулін у різних видів тварин має неоднакові деталі первинної структури, в основному амінокислотні залишки в положенні 8, 9, 10 (див. структуру білків.) Цей гормон може існувати у вигляді мономеру з відносною молекулярною масою 6000, а також у вигляді ди-, три-, тетра-, гекса- і октамерів. Утворення олігомерних форм інсуліну відбувається внаслідок взаємодії іонів цинку з імідазольними кільцями залишків гістидину поліпептидного ланцюга В.
Транспорт інсуліну до клітин-мішеней відбувається з участю а- і (З-глобулінів, які сполучаються з гормоном при виході із секреторних гранул. Біологічну активність має лише вільна форма інсуліну, яка утворюється при руйнуванні його комплексу з білками. Цей процес особливо інтенсивно відбувається в жировій тканині.
Головними клітинами-мішенями інсуліну є клітини печінки, м’язів і жирової тканини. Крім того, інсулін впливає на ряд інших клітин і тканин (фібробласти, лейкоцити, тканину молочної залози).
В основі молекулярних механізмів біологічної дії інсуліну, як і інших гормонів білкової природи, лежить його взаємодія з рецепторними білками, локалізованими на поверхні плазматичних мембран клітин-мішеней. Вважають, що рецептор інсуліну має глікопротеїдну природу, зокрема важливу роль у процесі рецепції відіграють залишки похідних нейрамінової кислоти — сіалові кислоти.
В організмі людини і тварин інсулін відіграє надзвичайно важливу роль. При недостатній секреції інсуліну (3-клітинами підшлункової залози виникає порушення багатьох ланок метаболічних процесів, які супроводжуються насамперед гіперглікемією (підвищення цукру в крові) і глюкозурією (поява цукру в сечі). Вони виникають внаслідок порушення транспорту глюкози в клітини і використання її тканинами організму. Вважають, що інсулін після фіксації його на поверхні клітин змінює їх так, що сприяє проникненню глюкози в клітини, де й відбувається її утилізація. Це так званий первинний ефект інсуліну. Усі інші ефекти інсуліну є, очевидно, лише наслідками, які залежать від первинного ефекту. Вторинними ефектами інсуліну є зниження вмісту цукру в крові, підвищення запасів глікогену в м’язах, посилення синтезу жирів і пептидів та ін.
Стимулюючи проникнення глюкози в клітини, інсулін тим самим зумовлює зниження вмісту її в крові І сприяє здійсненню процесів фосфорилювання глюкози під впливом глюкокінази і наступній утилізації її клітинами. Процес фосфорилювання глюкози має досить важливе значення, оскільки лише у фосфорильованій формі можливе наступне перетворення глюкози до кінцевих продуктів.
При порушенні внутрісекреторної функції підшлункової залози (її гіпофункції) виникає захворювання, що називається діабетом. Основні ознаки його: гіперглікемія, глюкозурія, спрага, підвищення апетиту. Втрата значної кількості цукру із сечею призводить до виснаження організму, слабкості, зниження працездатності і порушення зору. При цьому порушуються функції ряду внутрішніх органів — печінки, нирок, серцево-судинної і нервової систем.
Причини виникнення цього захворювання різні. Інколи хвороба виникає після психічних травм, інфекційних захворювань, надмірного харчування, при пухлинах і запальних процесах у підшлунковій залозі. Певне значення має також спадковий фактор. Більш як у 40 % хворих діабетом хворіють їхні батьки або близькі родичі.
При діабеті внаслідок порушення процесів утилізації глюкози і енергетичного голодування клітин зростає потреба в інших джерелах енергії, тому посилюється метаболізм білків і ліпідів. Спостерігається підвищення глюконеогенезу з білків і збільшення екскреції азоту із сечею.
Мобілізація ліпідів з жирових депо призводить до ліпемії, може виникнути жирове переродження печінки. Процеси розпаду жирів супроводжуються появою великої кількості недоокислених продуктів ліпідного обміну — кетонових тіл (β-оксимасляної і ацетооцтової кислот). При концентрації їх 20 моль/л виникає ацидоз — зміна pH крові. При ацидозі утруднюються процеси постачання глюкози до клітин центральної нервової системи, зменшується використання кисню тканиною мозку. В результаті всіх цих змін виникає стан, що називається діабетичною комою.
Від надмірного виділення інсуліну (гіперінсулінізм) внаслідок аденоми чи іншого захворювання підшлункової залози може виникнути гіпоглікемія. Це захворювання протилежне цукровому діабету. Його симптоми: слабкість, пітливість, почуття голоду, прискорений пульс, втрата свідомості. Такий стан може виникнути і внаслідок введення досить високих доз інсуліну при лікуванні цукрового діабету.
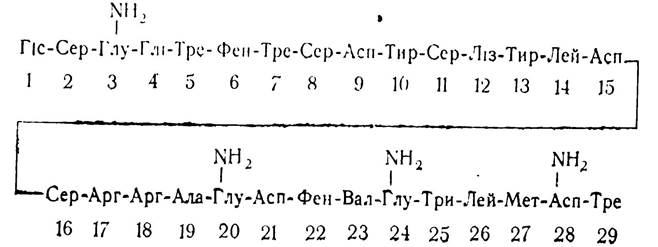
Глюкагон. Крім інсуліну, у підшлунковій залозі синтезується ще один гормон — глюкагон. Добуто глюкагон у кристалічному стані в 1953 р. Згодом було вивчено його хімічну природу. Виявилось, що глюкагон є низькомолекулярним пептидом, який складається з 29 залишків амінокислот та їх амідів.
Враховуючи специфічну дію глюкагону, його називають гіперглікемікоглікогенолітичним фактором (ГГФ). Подібно до інсуліну та інших поліпептидних гормонів, глюкагон у певних клітинах-мішенях зв’язується з глюкагоноспецифіч- ними рецепторними системами, що призводить до активації мембранної аденілатциклази і збільшення внутріклітинної цАМФ, внаслідок чого відбуваються активізація протеїнкіназ і прискорення процесів фосфорилювання специфічних білків. Крім того, глюкагон збільшує швидкість протеолізу білків у печінці.
У печінці глюкагон гальмує синтез жирних кислот, холестерину з ацетату і стимулює кетогенез, активує ліпази, підвищує кількість ацетил- і ацил-КоА. У нирках під впливом гормону підвищується клубочкова фільтрація, прискорюється рух крові, посилюється екскреція іонів натрію, хлору і сечової кислоти.
Глюкагон зумовлює збільшення глюкози в крові, стимулюючи процеси розпаду глікогену тільки в печінці, на відміну від адреналіну, який стимулює розпад глікогену в печінці і м’язах. Це пояснюється тим, що глюкагон стимулює перетворення неактивної фосфорилази печінки в активну форму в присутності відповідного ферменту, який відсутній у м’язах.
Гормони надниркових залоз
Надниркові залози розміщені безпосередньо над нирками, до яких вони тісно прилягають. Ці парні органи мають масу 7—12 г. Гормони, що продукуються в цих залозах, відіграють важливу роль у багатьох метаболічних процесах, забезпечують нормальне функціонування цілого ряду органів і систем організму. Ці залози є життєвонеобхідними і при видаленні їх піддослідні тварини гинуть протягом 2—3 днів.
Кожна надниркова залоза складається з двох структур — мозкової речовини і коркового шару, який становить 2/3 маси залози. Ембріональне походження, будова і гормони, що продукуються в цих структурах, різні, але в організмі між ними існує функціональна взаємодія.
Гормони мозкової частини надниркових залоз. У мозковій частині надниркових залоз продукуються два катехоламіни, які мають гормональну активність,—адреналін і норадреналін.
Крім адреналіну і норадреналіну, у мозковій частині надниркових залоз утворюється як проміжний продукт їх біосинтезу третій біологічно активний катехоламін — 3,4-діоксифенілетиламін (дофамін).
Адреналін і норадреналін впливають на деякі ланки обмінних процесів, особливо на процеси, пов’язані з енергетичним обміном. Характер біологічної дії обох катехоламінів однаковий, але норадреналін є більш ефективним медіатором, ніж адреналін.
Важливою функцією катехоламінів є здатність виявляти вазорепресорну дію. Зокрема, адреналін звужує кровоносні судини в шкірі і внутрішніх органах і розширює коронарні судини серця, підвищує частоту і силу серцевих скорочень, розслаблює гладеньку мускулатуру легень, бронхів та інших внутрішніх органів. Норадреналін виявляє дещо слабшу дію, зокрема, не сприяє розслабленню м’язів бронхів.
Гіпофункція мозкової частини надниркової залози зустрічається досить рідко. Явище гіперфункції встановлено при аденомах хромафінної тканини. При цьому виникають гіпертонія, глюкозурія, підвищення основного обміну.
Гормони кори надниркових залоз утворюють групу гормонів — кортикостероїдів, які регулюють значну кількість метаболічних процесів в організмі.
Діапазон дії цих гормонів досить широкий. Важко уявити такий аспект життєдіяльності організму, такий вид обміну речовин, які тією чи іншою мірою, безпосередньо чи опосередковано не зазнавали б впливу цих гормонів. У вивченні цієї групи гормонів велике значення мають дослідження радянського вченого О. О. Богомольця та його співробітників. Було вивчено хімічну природу, особливості структури кортикостероїдів та добуто перші препарати стероїдних гормонів. Академік О. О. Богомолець понад 60 років тому довів, що гормональну дію має секрет кори надниркових залоз — жироподібна речовина, що міститься в клітинах залози. Таким чином, вперше було доведено, що гормони кори надниркових залоз мають ліпідну природу. Нині з’ясовано структурну та хімічну природу великої групи стероїдних гормонів. Всього з кори надниркових залоз виділено понад 40 речовин, що мають стероїдну природу. Проте докладно вивчено лише частину цих речовин — кортикостерон, 11-дегідрокортикостерон, 17-оксикортикостерон, кортизон, гідрокортизон і альдостерон. Найважливішими з цих сполук, що постійно виділяються в кров і мають гормональну дію, є гідрокортизон, кортикостерон і альдостерон. Ці речовини — життєвонеобхідні. При введенні їх вдається продовжити життя тваринам з видаленою корою надниркових залоз. Інші речовини кори надниркових залоз мало впливають на метаболічні процеси, деякі з них е проміжними продуктами синтезу кортикостероїдів.
За хімічною будовою кортикостероїди подібні до холестерину, жовчних кислот і вітаміну D. Це похідні циклічного вуглеводню — стерану. За характером хімічної структури гормони кори надниркових залоз поділяють на кілька груп. Гормони першої групи містять 21 вуглецевий атом і є похідними вуглеводню прегнану. До цієї групи належать типові гормони надниркових залоз.
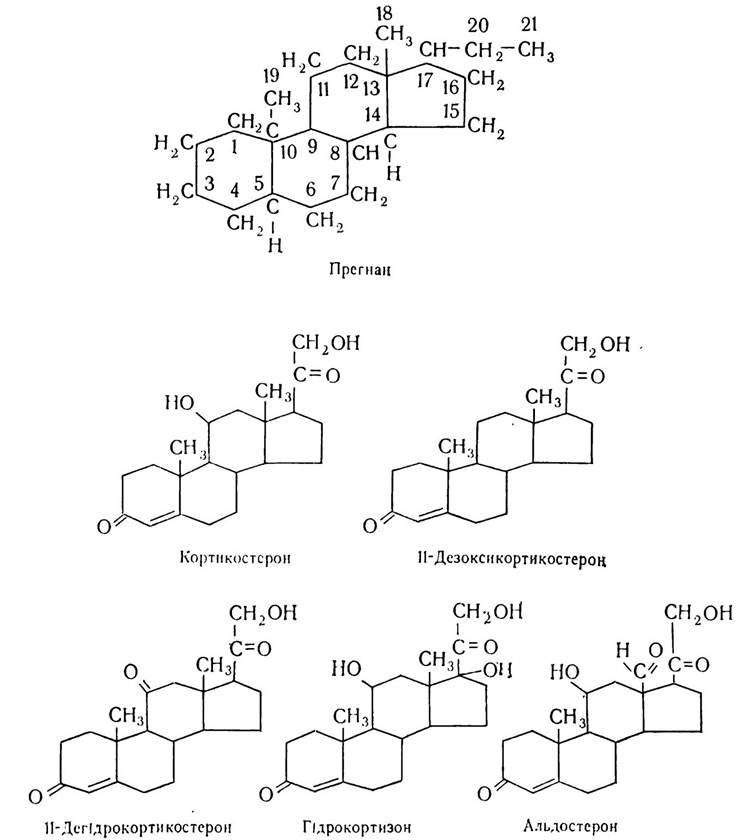
Крім того, тут синтезуються також стероїди, що містять 19 вуглецевих атомів і є похідними андростану.
Гістохімічна кора надниркових залоз поділяється на три зони. Зовнішня (клубочкова) зона продукує мінералокортикоїди, що регулюють обмін води та електролітів (альдостерон і 11-дезоксикортикостерон). Середня (пучкова зона) продукує глюкокортикоїди, які регулюють вуглеводний обмін (гідрокортизон, кортикостерон). Внутрішня (сітчаста зона) продукує гормони андрогени та естрогени, за структурою і функціями близькі до гормонів статевих залоз. Слід зазначити, що такий поділ функцій кортикостероїдів дещо умовний. Зокрема, гідрокортизон, який регулює в основному вуглеводний обмін, може деякою мірою виявляти й інші види активності. Однак вплив його на мінеральний обмін зовсім незначний. Альдостерон впливає в основному на мінеральний обмін, а вплив його на інші види метаболічних процесів становить 1/3 активності гідрокортизону. Кортикостерон менш активний, ніж гідрокортизон і альдостерон. Тобто гідрокортизон і альдостерон за характером реакцій, які вони викликають, займають крайнє положення, а кортикостерон — проміжне.
За добу в організмі людини синтезується 10—30 мг гідрокортизону (кортизону), 2—4 мг кортикостерону і 300— 400 мкг альдостерону. Отже, на гідрокортизон припадає 80 % загальної кількості кортикостероїдів. Він є основним гормоном кори надниркових залоз.
Навіть з неповного переліку біологічних функцій кортикостероїдів видно, що вони значно впливають на різні метаболічні процеси, зокрема, регулюють обмін білків, вуглеводів і ліпідів, води та електролітів, позитивно впливають на кровотворні процеси, підвищують резистентність організму проти несприятливих факторів.
Гормони гіпофізу
Гіпофіз — одна з найважливіших залоз внутрішньої секреції, яка впливає на різні метаболічні процеси, а також на формування функцій і структури організму. Враховуючи його функціональне значення, гіпофіз вважають своєрідним центром ендокринної системи.
Гіпофіз має еліпсоподібну форму, його маса 0,5—0,7 г. У структурі гіпофізу розрізняють три функціонально самостійних зони — передню (аденогіпофіз), середню (інтерме- діальна зона) і задню (нейрогіпофіз). Кожна з цих зон виділяє характерні для неї гормони, які регулюють ту чи іншу ланку обмінних процесів в організмі.
Вивчаючи функції гіпофізу, виявили, що після видалення цього органу у тварин порушуються функції (і навіть атрофуються) деяких залоз внутрішньої секреції — щитовидної, надниркових, статевих, а також ріст і цілий ряд обмінних процесів. А після пересадки гіпофізу гіпофізектомованим тваринам порушені функції відновлюються. Таким чином, було встановлено, що гіпофіз виділяє речовини, які регулюють функціональну діяльність ряду ендокринних залоз, впливають на обмінні процеси в організмі.
Пізніше було з’ясовано, що гормони гіпофізу не можуть компенсувати зміни, які виникають в організмі після видалення тієї чи іншої ендокринної залози, функціональна активність якої залежить від гіпофізу.
Отже, вплив гіпофізу на різні ланки обміну речовин виявляється не безпосередньо, а опосередковано, через вплив на функції інших ендокринних залоз, які перебувають під його регуляторною дією. Тому гормони гіпофізу називаються тропними гормонами (від грецького tropos — напрям).
За хімічною природою гормони гіпофізу є поліпептидами або білками з невеликою відносною молекулярною масою. У передній частці гіпофізу синтезуються гормони, серед яких найважливіше значення мають соматотропний гормон (СТГ), адренокортикотропний (АКТГ), тиреотропний (ТТГ), гонадотропні гормони (лютенізуючий, фолікулостимулюючий, пролактин), а також ліпотропіни.
Соматотропний гормон (гормон росту) виділили з екстрактів аденогіпофізу дослідники Г. Еванс і Дж. Лонг у 1921 р. Гормон має білкову природу і олігомерну структуру. Його відносна молекулярна маса 23 000— 46 000. Складається гормон росту з двох поліпептидних ланцюгів, які містять 191 амінокислотний залишок.
Характерною особливістю гормону росту є те, що він має високу видову специфічність. Хоч гормони деяких видів тварин і виявляють біологічну активність при введенні їх іншим тваринам, для людини біологічно активним є лише гормон приматів.
Різноплановий характер впливу соматотропного гормону на різні метаболічні процеси в організмі людини і тварин, на відміну від дії інших аденогіпофізарних гормонів, не зумовлений його впливом на інші ендокринні залози. При порушенні секреції соматотропного гормону виникає порушення деяких метаболічних процесів в організмі, зокрема, порушуються анаболічні процеси і процеси росту. При надмірному виділенні гормону (гіперфункції залози) в дитячому віці, до настання статевої зрілості і до завершення
процесів окостеніння, спостерігається посилений ріст органів і тканин організму, що призводить до гігантизму. Ріст індивідумів при цьому може становити 210—240 см. Надмірне виділення гормону росту в зрілому віці призводить до розвитку захворювання, яке називається акромегалією. Це захворювання виникає при гіперплазії та аденомах гіпофізу. Ознаками захворювання є непропорційне розростання окремих частин тіла, надмірний ріст кісток кінцівок, голови, обличчя, розростання м’яких тканин — носа, губ, підборіддя. Спостерігається надмірний ріст волосся на тілі, збільшуються внутрішні органи (спланхномегалія), спостерігається гіпертрофія ряду ендокринних залоз.
При недостатньому виділенні гормону росту (гіпосекреції) ріст сповільнюється, розвивається карликовість. Ріст при цьому не перевищує 90—120 см, відсутні вторинні статеві ознаки. Гіпофізарні карлики, на відміну від тиреогенних, не мають ознак деформації скелета і непропорційності різних частин тіла, хоч у них дещо збільшена голова. У розумовому розвитку вони не відрізняються від звичайних людей.
Специфічний вплив соматотропного гормону на процеси росту в організмі свідчить, що він відіграє важливу роль у забезпеченні анаболічних процесів, сприяє синтезу білків і нуклеїнових кислот, збільшенню рівня глюкози в крові (діабетогенний ефект), посилює транспорт сполук у клітини та їх засвоєння, стимулює ретикулоцитоз, хондро- і остеогенез.
Гормон росту впливає на тканини організму як безпосередньо, активацією мембранозв’язаної аденілатциклази, так і опосередковано, через інші агенти, зокрема сомато- медіни — вторинні ростостимулюючі фактори, виділені з різних органів — печінки, м’язів, нирок, крові.
Тиреотропний гормон (ТТГ) забезпечує нормальне функціонування щитовидної залози. При гіпофізек- томії у тварин спостерігається інволюція щитовидної залози з ознаками, що характерні для гіпотиреозу. Введення таким тваринам екстракту гіпофізу сприяє репаративним процесам і нормалізації функцій щитовидної залози.
За хімічною природою тиреотропний гормон є складним білком — глікопротеїдом з відносною молекулярною масою 28 000. Він має олігомерну будову і складається з двох субодиниць а- і р-, що зв’язані нековалентними зв’язками. Білкова частина гормону містить велику кількість сірковмісних амінокислот, які утворюють внутріланцюгові дисульфідні зв’язки, п’ять з яких стабілізують структуру α-субодиниці і шість — (3-субодиницІ молекули гормону. Чергування амінокислотних залишків в α-субодиниці гормону кожного виду тварин однакова, а первинна структура β-субодиниць у різних видів тварин різна. Тому вважають, що саме β-субодиниці забезпечують характерну гормональну активність тиреотропного гормону і регуляцію ним функцій щитовидної залози. Тиреотропний гормон впливає на метаболічні процеси в тканині щитовидної залози. Під впливом ТТГ стимулюються процеси вбирання йодидів з крові, включення йоду до складу тиреоїдних гормонів та вивільнення їх із залози активацією фолікулярних протео-' літичних ферментів, які розщеплюють білок-попередник. Крім того, тиреотропний гормон стимулює утворення цАМФ в тканині щитовидної залози, синтез РНК і білків, фосфогліцеридів і сфінголіпідів, здійснює транспорт і перетворення глюкози. Дія тиреотропного гормону на клітини щитовидної залози та інші клітини-мішені починається після зв’язування гормону із специфічними рецепторами плазматичних мембран, які мають глікопротеїдну природу. Вважають, що важливу роль у забезпеченні рецепторної функції відіграють сіалові кислоти, які є компонентами глікопротеїдів. Наступні процеси, що відбуваються після зв’язування гормону з рецепторами, є типовими для дії пептидних і білкових гормонів.
Адренокортикотропний гормон (АКТГ) стимулює ріст і функціональну активність кори надниркових залоз, регулює синтез і секрецію кортикостероїдних гормонів. За хімічною природою АКТГ є поліпептидом з відносною молекулярною масою 20 000. До його складу входять 39 залишків 15 різних амінокислот.
Синтезується АКТГ в аденогіпофізі у вигляді попередника, який має значно вищу відносну молекулярну масу і перетворюється в активну форму гормону внаслідок гідролітичного відщеплення частини поліпептидного ланцюга. Виявлено, що перші 24 амінокислотні залишки, розміщені біля N-кінця молекули, в усіх видів тварин ідентичні і забезпечують, очевидно, його біологічну активність. Інші амінокислотні залишки (25—39) в різних видів варіюють і визначають видову імунологічну специфічність гормону. Методом хімічного синтезу добуто поліпептид, що містить 24 амінокислотні залишки і має таку саму біологічну активність, як і природний гормон. У забезпеченні біологічної активності важливе значення мають залишки діаміномонокарбонових кислот — лізину та аргініну.
Основна роль АКТГ полягає в прискоренні синтезу кортикостероїдів у корі надниркових залоз з холестерину. Він може діяти як безпосередньо на тканини різних органів, так і опосередковано, за допомогою кори надниркових залоз.
При недостатній секреції АКТГ виявляються всі характерні симптоми гіпофункції кори надниркових залоз.
При гіперсекреції АКТГ виникає захворювання — гіпофізарний базофілізм (хвороба Кушінга). Для цього захворювання характерним є гіперглікемія, демінєралізація кісток, оволосіння обличчя, запалення сальних залоз.
В основі механізму дії АКТГ на клітини кори надниркових залоз лежить ряд взаємопов’язаних процесів, які стимулюються в клітинах після зв’язування АКТГ із специфічними рецепторами клітин-мішеней і стимуляції мемб- раннозв’язаної аденілатциклази.
Нейрогіпофіз ссавців виділяє гормони пептидної природи — окситоцин і вазопресин. Обидва гормони синтезуються в гіпоталамусі з одного білка-попередника (нейрофізину) і депонуються в нейрогіпофізі. У більшості нижчих хребетних знайдено гормон вазотоцин, який проявляє властивості обох гормонів.
За хімічною природою окситоцин і вазопресин є нонапептидами з відносною молекулярною масою відповідно 1082 і 1007. До їх складу входять шість залишків амінокислот і три аміди:
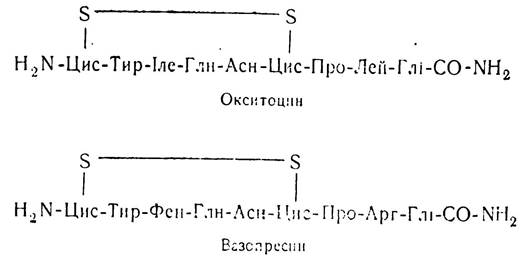
Як видно з наведених формул, окситоцин і вазопресин мають дуже подібні структури. Первинна структура гормонів відрізняється лише деякими деталями — вазопресин не містить в своєму складі ізолейцину і лейцину, а окситоцин — фенілаланіну та аргініну. Останнім часом синтезовано велику кількість аналогів окситоцину і вазопресину і вивчено їх біологічну активність. Функція гормонів нейрогіпофізу досить важлива для організму.
Вазопресин забезпечує діяльність однієї з найважливіших систем організму — системи регуляції водного обміну.
Нейрогіпофіз досить чітко реагує на зміну кількості води в організмі збільшенням або зменшенням секреції вазопресину. Вазопресин стимулює зворотне всмоктування води в дистальних ниркових канальцях і називається антидіуретичним гормоном.
Вважають, що важливою функцією гормону є активація ним ферменту гіалуронатліази, який розщеплює мукополісахарид-гіалуронову кислоту і збільшує проникність клітинних мембран, а отже, як результат — посилює реабсорбцію в ниркових канальцях. На клубочкову фільтрацію вазопресин не впливає.
При пошкодженні або атрофії нейрогіпофізу зменшується виділення гормону, внаслідок чого розвивається захворювання, що називається нецукровим діабетом. Хворі відчувають постійну спрагу, водночас значно підвищується виділення сечі. При важких формах захворювання людина випиває до 20 л води за добу і стільки ж виділяється сечі (поліурія).
Окситоцин вибірково підвищує тонус гладеньких м’язів матки і молочних залоз, сприяє посиленню їх скорочення. Секреція та активність окситоцину залежать від функціонального стану організму. Під час вагітності, наприклад, вони значно зменшуються, а під час родів збільшуються. У гінекологічній практиці гормон широко використовують для стимуляції родової діяльності.
Окситоцин сприяє також скороченню м’язів кишечника, сечового міхура, стимулює секрецію молока активацією гормону аденогіпофізу пролактину. На секрецію нейрогіпо- фізарних гормонів впливає ряд специфічних факторів. Це насамперед вплив центральної нервової системи, лікарських речовин.
Гормони гіпоталамусу
Гіпоталамус — основний орган гормональної регуляції, місце безпосередньої взаємодії нервової та ендокринної систем. Розміщений він у підбугровій ділянці проміжного мозку, яка одночасно є нервовим центром і ендокринним органом. У певних ділянках гіпоталамусу відбувається трансформація нервових імпульсів в ендокринний процес. Нейроендокринні центри гіпоталамусу безпосередньо зв’язані з різними ділянками мозку прямими і непрямими зв’язками, які здійснюються в основному через ретикулярну формацію і сіру речовину середнього мозку. Водночас гіпоталамус має безпосередні зв’язки з іншим важливим
ендокринним органом — гіпофізом, який контролює діяльність багатьох ендокринних залоз. Гіпоталамус і гіпофіз утворюють єдиний структурно-функціональний комплекс, який відіграє важливу роль у регуляції багатьох метаболічних процесів організму.
Гістохімічна будова гіпоталамусу неоднорідна. У великоклітинних ядрах переднього гіпоталамусу здійснюється синтез нейрогіпофізарних гормонів окситоцину і вазопресину, які транспортуються в нейрогіпофіз, де вони депонуються і використовуються в міру потреби.
У зоні дрібноклітинних ядер продукується група адсногіпофізарних рилізінг-факторів (від англ. to release — звільняти). Ці фактори відіграють провідну роль у регуляції функцій адено- і нейрогіпофізу. В ядрах гіпоталамусу синтезується сім стимуляторів (ліберинів) і три інгібітори І(статини) інкреції гіпофізарних гормонів. Частина цих факторів докладно вивчена, встановлено їх структуру і здійснено синтез.
Тіроліберин (стимулятор інкреції тиреотропного гормону) є трипептидом, що складається із залишків піроглутамінової кислоти, гістидину і проліну:
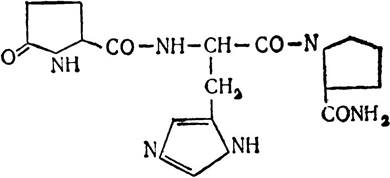
Вивчено ліберин лактогенного гормону, який є олігопептидом, і ліберин соматотропного гормону, який має полі- пептидну природу.
Виділено і вивчено також ряд статинів — соматостатин (інгібітор виділення соматотропного гормону), пролакто- статин (інгібітор секреції пролактину) та ін. Характерною особливістю рилізінг-факторів є їх висока специфічність відносно впливу на стимуляцію чи гальмування виділення в кров певного гормону.
Гормони рослин
Як в організмі людини і тварин, так і в рослинних організмах здійснюється чітка регуляція метаболічних процесів на внутріклітинному та міжклітинному рівнях. Вивченню природи факторів, що визначають міжклітинну взаємодію в рослинних організмах, приділяється велика увага, бо створення ефективної системи керування ростом і розвитком рослин є одним з найважливіших завдань, які стоять перед вченими в галузі рослинництва.
Широке дослідження способів взаємодії клітин, тканин і органів рослин, процесів їх росту та розвитку дало змогу встановити хімічну природу сигналів, які забезпечують ці процеси. До таких ендогенних факторів регуляції росту і розвитку рослинних організмів належать насамперед фіто- гормони — речовини, які утворюються в тканинах рослин.
У рослинних організмах фітогормони виконують різноманітні функції. Вони регулюють процеси росту, розвитку і диференціювання органів і тканин, перехід від однієї фази розвитку до іншої, формування статі квіток, забезпечення цвітіння і плодоношення, перебування їх в стані спокою чи вихід з нього. Гормони регулюють усі фази життєдіяльності рослин. Зв’язок між окремими органами і тканинами рослинного організму здійснюється з участю їх циркуляційної системи, яка забезпечує транспорт різних речовин від кореневої системи вгору по ксилемі та від листків вниз по флоемі. Крім того, відбувається активне транспортування через клітинні мембрани проти градієнта концентрації. Таким чином, фітогормони, утворені в одних органах і тканинах, потрапляють в інші, де спрямовують і стимулюють певні метаболічні процеси, забезпечуючи функціональну цілісність рослинного організму, узгоджуючи діяльність усіх його систем. Відомо такі групи речовин, які належать до категорії фітогормонів: ауксини, гібереліни, цитокініни, абсцизова кислота та етилен.
Ауксини (стимулятори росту). Відкриття ауксинів пов’язане з вивченням явища фототропізму в колеоптилях вівса. Встановлено, що проростки рослин, які вийшли на поверхню, завжди згинаються в бік джерела світла, наближаючи до нього свої фотосинтезуючі органи. Причому, час від початку дії подразника до видимого ефекту становить 25—60 хв. Затемнюючи різні ділянки колеоптилю, дослідили, що чутливість до світла має лише його верхівка (~ 250 мкм). Нижче розміщена зона згину, де спостерігається нерівномірний ріст клітин.
Отже, верхівка колеоптилю містить якийсь носій інформації про напрям світла, а переміщення цього носія визначає фототропічний згин колеоптилю.
Роль ауксинів в явищах фототропізму було висвітлено в гормональній теорії американського дослідника Ф. Вен- та. Ауксин, що утворюється в верхівці колеоптилю, транспортується вниз, причому концентрація його на затемненому боці рослини значно більша, ніж на освітленому, внаслідок чого забезпечується більш інтенсивний ступінь росту затемненого боку рослини і повертання її в бік джерела світла. Особливо добре спостерігати явища фототропізму в такої рослини, як соняшник.
Ауксини містяться у верхівках стебел не тільки вищих рослин, а й у водоростях, грибах, бактеріях. Часто рослини одержують ауксини від своїх паразитів або симбіонтів. Так, утворення бульбочок на коренях бобових рослин відбувається під впливом ауксинів бульбочкових бактерій. Утворення різних наростів на рослинах зумовлене ураженням їх патогенними мікроорганізмами, що виділяють ауксини.
Поширюються ауксини вздовж рослини від місця їх утворення — верхівки стебла до зони росту кореня. З їх участю стимулюється ріст стебла і гальмується ріст бокових коренів.
Вивчаючи хімічну природу ауксинів, виявили аналогічні речовини в сечі, фекаліях, дріжджах. Виявилось, що діючою речовиною ауксинів є індолілоцтова кислота, яка утворюється в результаті ряду перетворень з амінокислоти триптофану:
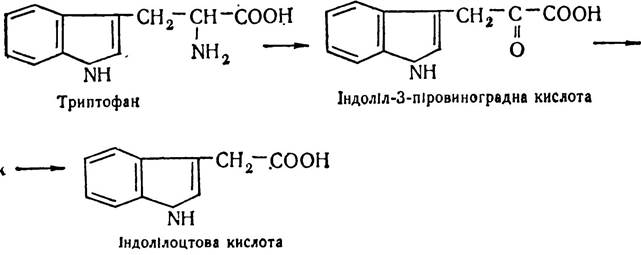
Індолілоцтова кислота регулює як поділ, так і ріст клітин, впливає на багато процесів, що відбуваються в клітині і позаклітинному середовищі. Зокрема, ауксини беруть участь у регуляції процесів цвітіння, запліднення, росту пагонів, регулюють процеси опадання листя, зав’язі і плодів. Опадання пов’язане з формуванням в черешках чи плодоніжках спеціалізованого (відокремлюючого) шару, який виділяє ферменти, що руйнують зв’язки між окремими клітинами. А провідні судини закриваються пробками з полісахаридів. Ці процеси пов’язані із зменшенням концентрації ауксинів у певних частинах рослин. Введення додаткової кількості ауксинів запобігає цьому.
Дія ауксинів залежить від їх концентрації. Порогові концентрації стимулюють процеси росту, а концентрації, вищі від порогових, гальмують їх. Тому в рослинах існує система регуляції вмісту ауксину і підтримання концентрації його нижче від рівня, який виявляє гальмівну дію. Цей процес здійснюється руйнуванням ауксину або зв’язуванням його з молекулами інших речовин з утворенням неактивних комплексів. У такому вигляді фітогормон може циркулювати в рослині, не впливаючи на тканини, з якими він перебуває в постійному контакті, тобто утворюється своєрідний запас (депо) речовин-регуляторів.
Молекулярні механізми біологічного впливу ауксинів ще не повністю з’ясовані, припускають, що вони (подібно до інших гормонів) стимулюють процеси транскрипції.
Гібереліни. Відкриття гіберелінів пов’язане з дослідженням японським ученим Є. Куросава причин виникнення захворювання рису. Ознакою цього захворювання був надмірний ріст вражених хворобою рослин. Виявили, що причиною цього захворювання є патогенний гриб гіберела, який виділяє в тканини рослин біологічно активні речовини, які було названо гіберелінами. Через деякий час з фітопатогенного гриба виділили в кристалічному стані речовину, яка стимулювала ріст рослин. Пізніше гібереліни були виділені з тканин багатьох рослин, при цьому виявилось, що вони є нормальними продуктами їх життєдіяльності.
За хімічною природою гібереліни є тетрациклічними карбоновими кислотами, які належать до класу дитерпенів, сполук досить поширених у природі. Вони входять до складу камфори, скипидару, різних смолистих речовин.
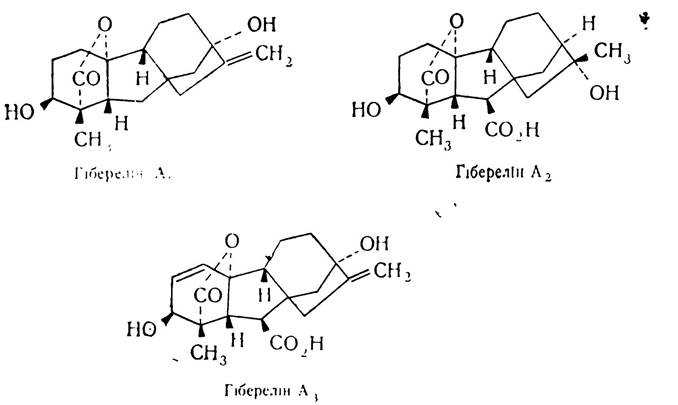
У наш час з рослинних тканин виділено понад 50 гіберелінів. Найпоширенішим є гіберелін А3 (гіберелова кислота). Гібереліни, як і ауксини, впливають на процеси росту і розвитку рослин. Стимуляція росту пов’язана з прискоренням процесів поділу, хоч важливим є також ефект розтягування клітин. Особливо інтенсивно під впливом гіберелінів відбувається ріст стебел як трав’янистих, так і деревовидних рослин. Крім того, під дією гіберелінів збільшуються розміри листків, квіток, суцвіть, плодів. Однак, найважливішим є те, що вони забезпечують процеси цвітіння і проростання, тобто перехід рослин з однієї фази розвитку в іншу.
Ще в 1880 р. німецький ботанік Ю. Сакс, підтримуючи гіпотезу Ч. Дарвіна про речовини — стимулятори росту, висловив припущення про існування формоутворюючих речовин. Однак тривалий час воно не було підтверджене. Лише в 30-х роках XX ст. радянськими фізіологами було розроблено гормональну теорію цвітіння. У 60-х роках, вивчаючи вплив гіберелінів на процеси цвітіння, встановили, що саме вони утворюються в рослинах при тривалому освітленні і знижених температурах та індукують процеси цвітіння. Гібереліни утворюються в насінні, листках, верхівкових бруньках, корінні. Вміст їх в рослинах становить 1·10-8—1·10—6 % від маси сухої речовини. Насіння плодів, що ростуть, виділяє гібереліни в плодове тіло, при видаленні насіння ріст плодів припиняється.
Гібереліни відіграють роль хімічних факторів, які стимулюють вихід рослин зі стану спокою. Механізм впливу гіберелінів на біохімічні процеси в організмі рослин з’ясовано не повністю. Вважають, що вони впливають на них через ферментні системи, які беруть участь у гідролітичному розщепленні резервних полісахаридів (крохмалю) до моносахаридів — основного енергетичного матеріалу клітин. На початковій стадії проростання зародок виділяє гібереліни, які індукують синтез а-амілази і гідролітичне розщеплення полісахаридів до моносахаридів, необхідних для росту зародка. Одна молекула гібереліну індукує утворення 2000 молекул амілази.
Цитокініни. У 30-х роках було виділено із спиртового екстракту квасолі речовину, яка стимулювала поділ клітин. Згодом речовини з подібними властивостями було виділено з плодів і насіння багатьох рослин. Особливо високий вміст їх виявився в насінні кукурудзи в фазі молочно-воскової стиглості. Із 70 кг такого насіння було виділено 1 мг речовини, яку назвали зеатином. Враховуючи специфічний вплив цих речовин на процеси росту і поділу клітин, їх було названо цитокінінами (цитос — клітина, кінез — поділ). Основна функція цитокінінів — забезпечення взаємного впливу і зв’язку між кореневою системою та наземною частиною рослини. У 1949 р. радянський фізіолог Д. А. Сабінін висловив думку про те, що в корінні рослин синтезується фітогормон, який регулює ріст стебла і листя. Це припущення грунтувалось на узгодженості процесів росту і розвитку надземної частини рослин зі станом їх кореневої системи та її можливістю забезпечити наземну частину потрібними речовинами, що є необхідною умовою нормального функціонування і виживання рослин у несприятливих умовах. Отже, у корінні повинна бути система регуляції росту надземної частини незалежно від умов живлення рослин. Виявилось, що коренева система регулює не тільки ріст надземної частини, а й кількість активно функціонуючого та відмираючого листя, бо зберігання правильної пропорції між ними є одним з важливих факторів гармонійного розвитку рослин. Крім того, забезпечуючи зв’язок між кореневою і надземною частинами рослин, цитокініни не тільки регулюють надходження поживних речовин і стимулюють синтез білка, а й посилюють основну функцію зелених частин рослини — фотосинтез.
З участю цитокінінів підвищується стійкість рослин про- . ти несприятливих факторів — зневоднення, хімічних агентів, опромінення, інфекційних захворювань, високих температур.
Вплив цитокінінів на рослинні організми досить різноманітний. Особливо активно стимулюють цитокініни поділ клітин у точках росту стебел і в зародках насіння, що проростає. Цитокініни мають здатність знімати ефект епікального домінування. Коли цитокініни нанести на точку росту стебла, то можна викликати передачу ролі основного пагона одному з бокових або зумовити одночасний ріст кількох бокових пагонів. Точка росту, що дістала додаткові цитокініни, починає посилено рости і притягувати поживні речовини. Здатність цитокінінів викликати надходження поживних речовин до різних ділянок рослин використовується різними паразитами рослин і організмами, які живуть з ними в симбіозі. Синтезуючи цитокініни і вводячи їх в рослину, вони забезпечують приплив поживних речовин до місця свого знаходження.
Найпоширенішими цитокінінами є зеатин, кінетин і N, N'-дифенілсечовина.
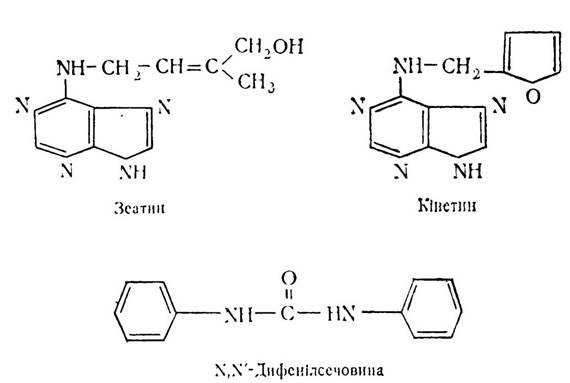
Механізм впливу цитокінінів на клітини-мішені ще не повністю з’ясовано. Вважають, що вони впливають на мембрани клітин, змінюючи проникність для різних іонів, а також посилюють процеси синтезу в клітинах внаслідок надходження поживних речовин. Підвищення синтезу РНК і білків в клітинах збільшує кількість рибосом та іРНК, тРНК, тобто розміри синтетичного апарату та його активність, можливо, активацією ферменту РНК-полімерази.
Етилен. Біологічну роль етилену вивчали на основі дослідження процесів дозрівання плодів і опадання листя рослин. Ці процеси є результатом фізіологічних змін, які призводять до старіння органів і організму в цілому, Вони супроводжуються зниженням поділу рослинних клітин, послабленням метаболічних і фотосинтетичних процесів. Було встановлено, що перехід у фазу старіння є гормональним процесом, який починається в листках після досягнення ними максимальних розмірів, а в рослин у цілому — в період закладання органів розмноження. Саме тканини, що старіють, досить чутливі до дії етилену. Надходження етилену з листків у черешки є сигналом до їх опадання. Пошкодження листкових пластинок зумовлює активне виділення етилену і опадання цих листків.
За хімічною природою етилен є ненасиченим газоподібним вуглеводнем. За агрегатним станом він відрізняється від інших фітогормонів. Синтез його в рослинах здійснюється з участю амінокислоти метіоніну:
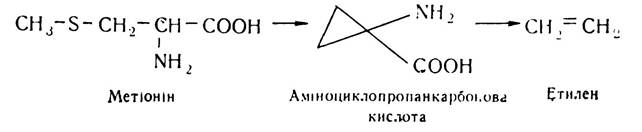
Аміноциклопропанкарбонова кислота легко відщеплює етилен і є, очевидно, транспортною формою його в організмі рослин. Для перетворення аміноциклопропанкарбонової кислоти в етилен необхідний кисень, тому створенням анаеробних умов можна затримати процеси дозрівання плодів.
Велика кількість етилену міститься в органах рослин, що старіють, у листках перед опаданням, зів’ялих квітках, дозріваючих плодах. Гормональні функції етилену досить різноманітні. Крім стимуляції процесів старіння, він регулює процеси дозрівання плодів, формування їх забарвлення. Дозрівання плодів завжди супроводжується підвищенням концентрації в них етилену. За допомогою етилену можна стимулювати цвітіння деяких рослин, змінювати співвідношення тичинкових і маточкових квіток, зумовлювати проростання насіння, виводячи його зі стану спокою.
Абсцизова кислота. У 50-х роках з екстрактів рослинних тканин було виділено речовину, яка пригнічувала ріст, зумовлювала опадання листя та стимулювала стан спокою в рослин. Враховуючи характер дії цієї речовини, її було названо абсцизіном (що в перекладі з англійської означає опадання). Пізніше в найбільш активної з цих речовин було виявлено кислотні властивості і названо абсцизовою кислотою (АБК).
Фізіологічна роль абсцизової кислоти різноманітна — вона пригнічує проростання коренеплодів, насіння різних, рослин, прискорює процеси опадання листя і старіння, пригнічує процеси фотосинтезу. АБК гальмує не тільки ріст, а й енергозабезпечення клітин, зокрема знижує проникність мембран і постачання клітин неорганічним фосфатом, що гальмує процеси синтезу макроергічних сполук. Абсцизова кислота регулює також стан продихів рослин, через які відбуваються дихання і транспірація. Якщо рослина втрачає 10 % води, концентрація АБК у листках зростає в сотні разів за короткий час, що призводить до закриття продихів. Гальмування процесів життєдіяльності з участю АБК не впливає на рослини негативно, бо фізіологічна роль її залежить від концентрації в рослинних тканинах. При зниженні концентрації гальмівний вплив її знімається. Крім того, для нормального проходження процесів життєдіяльності в рослинних організмах потрібно забезпечити регулюючий ефект внаслідок дії на різні метаболічні процеси. Для цього потрібно вдатись до комплексної дії різних фі- тогормонів, одні з яких стимулюють, а інші — гальмують певні ланки обміну речовин. Так, у процесі утворення, росту і дозрівання плодів спостерігається як одночасна, так і послідовна дія різних гормонів. У процесі формування стиглого плода беруть участь усі відомі групи фітогормонів. В утворенні зав’язі, рості плодового тіла беруть участь гормони росту, а при дозріванні та опаданні плодів важливе значення має дія етилену і абсцизової кислоти. Комплексною дією різних гормонів регулюються і інші процеси життєдіяльності рослинних організмів.
Використання гормонів у медицині та сільському господарстві
У наш час гормональні препарати широко використовують у медицині для нормалізації обмінних процесів в організмі. їх можна поділити на три групи. До першої групи належать екстрактивні препарати натуральних гормонів, які добувають безпосередньо із свіжих ендокринних залоз тварин. У чистому кристалічному стані з органів тварин добувають інсулін, адреналін, естрон та інші гормони. Гормони, які не можна добути в чистому стані, використовують у вигляді екстрактів, що являють собою суміші гормональних речовин з баластними домішками. Це, зокрема, кортин, паратиреоїдин, пітуїтрин, а також гормональний препарат з висушеної щитовидної залози — тиреоїдин.
Другу групу гормональних препаратів становлять синтетичні аналоги стероїдних і статевих гормонів, гормони кори надниркових залоз та ін. Із здійсненням хімічного синтезу гормональних препаратів можливості гормонотерапії значно розширились. Важливість цього стає зрозумілою, коли врахувати, що кількість гормонів у біоматеріалі — ендокринних залозах тварин, з яких їх виділяють, дуже мала. Налагодження промислового виробництва гормональних препаратів на основі їх хімічного синтезу відкриває великі можливості для лікування багатьох небезпечних захворювань. Крім того, синтетичні аналоги гормонів у багатьох випадках мають набагато вищу біологічну активність, а тому можна використовувати менші дози їх для лікування і профілактики різних захворювань, а це запобігає небажаній побічній дії на організм гормонів.
Третю групу гормональних препаратів становлять замінники гормонів, які відрізняються від них деякими деталями в хімічній будові, але мають аналогічну гормональну дію. Це, зокрема, групи естрогенних, андрогенних, кортикоїдних, гонадотропних та інших препаратів.
Гормональні препарати особливо широко використовують для лікування ендокринних захворювань, а також різних порушень обміну речовин. Гормонотерапія грунтується на використанні фармакодинамічних властивостей гормональних препаратів для впливу на патологічні процеси зміною реактивності організму, регулюванням обмінних процесів і ліквідацією симптомів захворювання. При лікуванні ендокринних захворювань, залежно від виду патології, застосовують компенсаторну, стимулюючу та блокуючу гормонотерапію.
Компенсаторну терапію застосовують у разі випадання або частковій гіпофункції залоз внутрішньої секреції. Цукровий діабет лікують інсуліном, адісонову хворобу—препартом кори надниркових залоз, тетанію — паратгормоном. Введення гормонів при цьому зумовлює усунення порушень, але не впливає на їх причину, тобто на функцію відповідної залози, і тому має тимчасовий характер.
Стимулююча гормонотерапія застосовується для ліквідації гіпофункцій залоз препаратами тропних гормонів аденогіпофізу. Блокуюча терапія — при деяких ендокринних захворюваннях, що пов’язані з гіперфункцією певних залоз.
Гормональні препарати використовують не тільки для лікування ендокринних захворювань. Наприклад, гормоном інсуліном лікують різні токсикози, психічні захворювання, хвороби внутрішніх органів тощо.
Гормональні препарати використовують не лише в медицині, а й у тваринництві для лікування або біостимулювання того чи іншого виду обміну речовин для підвищення продуктивності тварин. Використання гормонів як добавок до харчових раціонів не завжди раціональне, оскільки багато гормонів при цьому втрачає свою біологічну активність. Здебільшого їх вводять у вигляді ін’єкцій і методом імплантації ендокринних залоз під шкіру. За допомогою ін’єкцій інсуліну і гормону росту дістають приріст маси 5—6-місячних поросят на 10—18 %. Введенням гормону щитовидної залози збільшують настриг вовни з ягнят.
Для підвищення продуктивності тваринництва використовують також синтетичні антитиреоїдні препарати (гойтрогени) типу тіоурацилу — метилтіоурацили. Якщо в птахівництві використовувати їх по 0,03 г на 100 г комбікорму протягом 14—16 днів, то продуктивність птиці підвищиться більш як на 30 %.
У рослинництві широко використовуються фітогормони — речовини, які регулюють складні процеси життєдіяльності рослинних організмів. Використання фітогормонів для потреб сільського господарства стало можливим лише після докладного вивчення і встановлення їх хімічної структури та синтезу речовин, які діяли аналогічно на метаболічні процеси в організмі рослин, тобто коли фітогормони стали доступними для широкого використання в практиці рослинництва.
Серед таких речовин найбільш поширеними є стимулятори (ауксини, гібереліни), сповільнювачі росту (ретарданти), гербіциди, дефоліанти та інші речовини. Зараз в практиці рослинництва широко використовуються похідні феноксіоцтової кислоти:
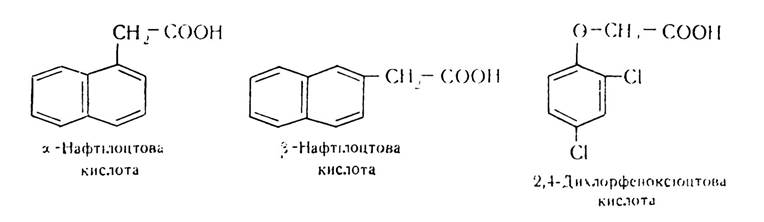
Більшість цих речовин є або аналогами фітогормонів за характером дії на рослини, або так чи інакше впливають на гормональний баланс рослин, регулюючи певні ланки метаболізму. Ці речовини в певних дозах можуть стимулювати або гальмувати процеси росту рослин. У першому випадку вони діють як ауксини чи гібереліни, у другому — як гербіциди.
Синтетичні регулятори застосовуються в різних галузях сільського господарства. Досить перспективним є використання їх для керування станом спокою рослин. Так, обробка картоплі розчинами гіберелінів і тіосечовини через кілька днів стимулює проростання бульб. З цією метою використовують регулятор росту етрел. Його застосовують також і для боротьби з бур’янами, насіння яких може довго зберігатися в грунті, не проростаючи, тому не чутливе до дії гербіцидів. Внесення в грунт 0,01—1 кг/га етрелу стимулює проростання насіння бур’янів, які потім легко знищують гербіцидами.
Регулятори росту рослин використовують і для підвищення врожайності ряду сільськогосподарських культур. Так, обробка гіберелінами винограду (дозою 30 г/га) зумовлює розпушення грон і збільшення розмірів ягід. Прибавка врожаю при цьому становить 50—100 %.
Серед проблем рослинництва найважливішою є підвищення стійкості високопродуктивних сортів різних культур проти несприятливих умов — підвищеної і зниженої температури, недостачі або надлишку вологи та ін. Причому селекційні методи підвищення продуктивності, як правило, знижують ці показники. Тому в наш час широко використовують речовини, які мають антигіберелінову активність — ретарданти, що блокують систему росту і розтягування стебла з участю гіберелінів. Ці речовини зумовлюють вкорочення і потовщення стебел рослин, розширення листкових пластинок, підвищення інтенсивності зеленого забарвлення їх, сприяють росту кореневої системи і не впливають на органи плодоношення. Найпоширенішими ретардантами є похідні нікотину, четвертинні амонійні солі, похідні янтарної кислоти та ін. Деякі з них використовують для боротьби з поляганням злакових культур — озимої пшениці, жита та овочів у теплицях, щоб не витягувались при недостачі світла.
Диметилгідразид янтарної кислоти є діючою речовиною зарубіжного ретарданту «алар» та вітчизняного «нора». Використовують їх в садівництві для стимулювання закладання квіткових бруньок. Яблуні обробляють 0,2—0,5-процентним аларом через 2 тижні після цвітіння, що дає змогу прискорити плодоношення молодих садів.
Велику роль відіграють регулятори росту в підвищенні ефективності фотосинтезу. У звичайних умовах лише 1 % світлової енергії, що падає на листок, перетворюється в хімічну. У польових умовах ефективність цього процесу обмежується здатністю рослин фіксувати оксид вуглецю (II), Наприклад, 1 га кукурудзи в період росту повинен асимілювати 22 000 кг СО2 і переробити 100 000 т повітря. Для підвищення ефективності цього процесу треба змінити форму наземної частини рослин так, щоб поліпшити циркуляцію повітря в посівах, а також змінити ступінь розкриття продихів, через які СО2 потрапляє в листки.
Широко використовуються в рослинництві дефоліанти — речовини, які сприяють опаданню листя. Найбільш поширеними з них є синтетичні похідні дифенілсечовини, в яких фенільні фрагменти замінені на гетероциклічні групи. Це так звані гормональні гербіциди, які в малих дозах виявляють гормональну дію, а в великих — токсичну. Так, дефоліант тіадіазурон широко застосовують для видалення листя на плантаціях бавовнику. Дефоліація бавовнику — необхідний етап, що передує збиранню врожаю, щоб запобігти забрудненню листям цінної сировини.
У рослинництві використовуються також речовини, які регулюють процеси цвітіння і плодоношення. Ці речовини забезпечують одночасне раннє дозрівання плодів, що дає змогу організувати машинне збирання їх і запобігти втратам врожаю. Для цього використовують продуценти етилену.
Для регулювання плодоношення використовують також ауксини. Помідори в період цвітіння при несприятливих умовах втрачають зав’язі, і при цьому утворюються дрібні плоди, що пов’язано з недостачею ендогенних ауксинів. Обробка їх ауксинами, зокрема 4-хлорфеноксіоцтовою кислотою, сприяє кращому плодоутворенню. Для цілорічного вирощування цитрусових перед збиранням врожаю їх обробляють 0,0008-процентним розчином 2,4-дихлорфеноксі- оцтової кислоти, внаслідок чого вони тривалий час зберігаються на деревах зеленими.
Регулятори процесів росту використовують також для зберігання сільськогосподарської продукції. Зокрема, у картоплесховищах з метою запобігання проростанню картоплі використовують метиловий ефір α-нафтилоцтової кислоти дозою 3 кг на 1 т. Запобігає проростанню картоплі і гідразид малеїнової кислоти (ГМК). Цим регулятором дозою 2,5 кг на 1 га за 15 днів до збирання обробляють надземну частину картоплі. Регулятор проникає в бульби, нагромаджується там і запобігає проростанню.
Необхідно зазначити, що більшість регуляторів росту малотоксичні для людини, тварин і самих рослин. Таким чином, в арсеналі рослинництва є велика кількість речовин, які дають змогу спрямовувати метаболічні процеси в організмі рослин в бажаному для людини напрямі.