БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 7. ЕНЗИМОЛОГІЯ
7.3. Кофактори й коферменти
7.3.1.Кофактори
Понад 25 % усіх ферментів для прояву повної каталітичної активності потребують наявності іонів металів. Розглянемо роль кофакторів у ферментативному каталізі.
Іони металу виконують функцію стабілізаторів молекули субстрату, активного центру ферменту й конформації білкової молекули ферменту, а саме третинної та четвертинної структур.
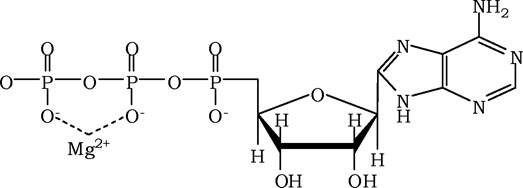
Іони металів - стабілізатори молекули субстрату. Для деяких ферментів субстратом слугує комплекс перетворюваної речовини з іоном металу. Так, для більшості кіназ одним із субстратів виступає не молекула АТФ, а комплекс Mg^-АТФ. У цьому випадку іон Mg2+ не взаємодіє безпосередньо з ферментом, а
задіяний у стабілізації молекули АТФ і нейтралізації від'ємного заряду субстрату, що полегшує його приєднання до активного центру ферменту:
Схематично роль кофактора під час взаємодії ферменту й субстрату можна зобразити як комплекс E-S-Me, де Е - фермент, S - субстрат, Ме - іон металу. Як приклад можна навести розміщення субстратів в активному центрі гексокінази (рис. 7.3). Гексокіназа каталізує перенесення кінцевого, γ-фосфатного залишку молекули АТФ на глюкозу з утворенням глюкозо-6-фосфату:
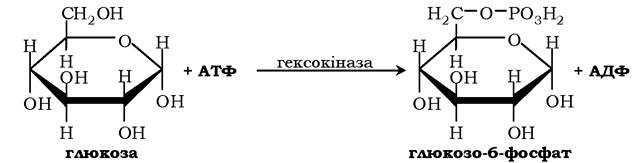
Іон Mg2+бере участь у приєднанні та "правильній" орієнтації молекули АТФ у активному центрі ферменту, послаблюючи фосфоефірний зв'язок і полегшуючи перенесення фосфату на глюкозу.
Іони металу - стабілізатори активного центру ферменту.
Іноді іони металу слугують "містком" між ферментом і субстратом. Вони виконують функцію стабілізаторів активного центру, полегшуючи приєднання до нього субстрату й перебіг хімічної реакції. У деяких випадках іон металу може сприяти приєднанню коферменту. Указані функції виконують такі метали, як Mg2+, Mn2+, Zn2+, Co2+, Mo2+. У разі відсутності металу ці ферменти не активні. Вони отримали назву "металоензими". Схематично даний процес взаємодії ферменту, субстрату й металу можна зобразити наступним чином: Е-Ме-S. До металоензимів належить фермент піруваткіназа (рис. 7.4), який каталізує реакцію:
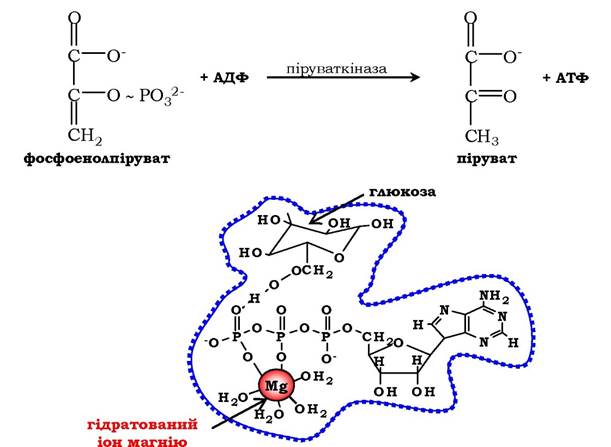
Рис. 7.3. Участь іонів магнію в приєднанні субстрату в активному центрі гексокінази.
В активному центрі гексокінази є ділянки зв'язування для молекули глюкози і комплексу Mg2+-АТФ. У результаті ферментативної реакції відбувається перенесення кінцевого γ-фосфатного залишку молекули АТФ на глюкозу з утворенням глюкозо-6-фосфату
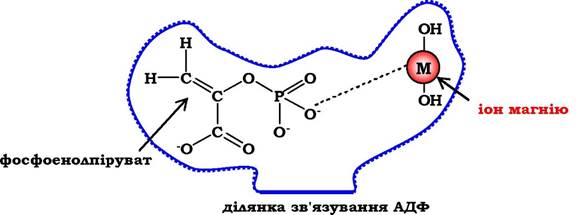
Рис. 7.4. Участь іонів магнію в приєднанні субстрату в активному центрі піруваткінази:
активний центр піруваткінази має ділянки зв'язування для фосфоенолпірувату й АДФ. Mg2+ бере участь у стабілізації активного центру, що полегшує приєднання фосфоенолпірувату. Унаслідок ферментативної реакції утворюється піруват і АТФ
Іони металів забезпечують зберігання третинної, четвертинної структури молекули ферменту. Такі ферменти за відсутності іонів металів здатні до хімічного каталізу, але вони нестабільні. Їхня активність знижується й навіть повністю зникає внаслідок невеликих змін рН, температури та інших незначних змін зовнішнього середовища. Отже, іони металів виконують функцію стабілізаторів оптимальної конформації білкової молекули.
Іноді в стабілізації третинної структури беруть участь іони лужних і лужноземельних металів. Так, для підтримання третинної конформації піруваткінази потрібні іони K+. Для стабілізації четвертинної структури алкогольдегідрогенази, яка каталізує реакцію окиснення етанолу, необхідні іони цинку. Алкогольдегідрогеназа складається з чотирьох субодиниць з молекулярною масою 151 кДа. До складу ферменту входять чотири атоми Zn2+. Видалення Zn2+ приводить до втрати ферментом активності за рахунок дисоціації на чотири неактивні субодиниці з молекулярною масою 36 кДа (рис. 7.5).
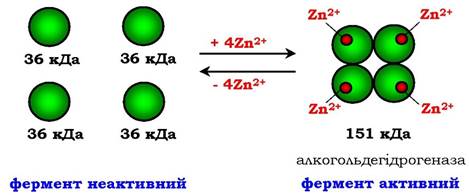
Рис. 7.5. Стабілізація четвертинної структури алкогольдегідрогенази іонами цинку
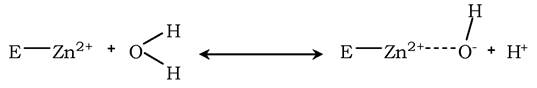
Роль металів у ферментативному каталізі. Не менш важливою є роль іонів металів у здійсненні ферментативного каталізу. Під час електрофільного каталзу вони часто беруть участь у стабілізації проміжних сполук. Найчастіше цю функцію виконують іони металів зі змінною валентністю, які мають вільну d-орбіталь і виступають як електрофіли. Насамперед, це такі метали, як Zn2+, Fe2+, Mn2+, Cu2+. Іони лужних металів, такі як Na+ і K+, не мають цієї властивості. Як приклад можна розглянути функціонування ферменту карбоангідрази - цинковмісного ферменту, що каталізує реакцію утворення вугільної кислоти:
![]()
Іон Zn2+ у результаті гідрофільної атаки цинк-ензиму бере участь в утворенні Н+ і ОН+ іонів із молекули води:
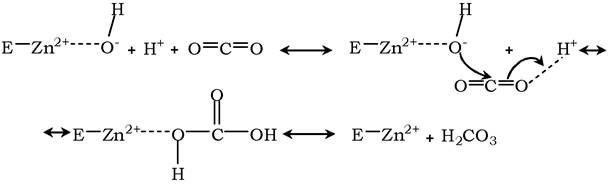
Протон і гідроксильна група послідовно приєднуються до діоксиду вуглецю з утворенням вугільної кислоти:
Іони металів зі змінною валентністю можуть також брати участь у перенесенні електронів. Наприклад, у цитохромах (гемовмісних білках) іон заліза здатен приєднувати й віддавати один електрон:
![]()
Завдяки цій властивості, цитохроми беруть участь в окисно- відновних реакціях.
Інший приклад участі іонів металів в окисно-відновних реакціях - робота ферменту дофамінгідроксилази, що каталізує реакцію утворення норадреналіну за участю вітаміну С. За окисно-відновні властивості дофамінгідроксилази відповідає іон міді (рис. 7.6). Фрагмент, який містить іон Cu2+, не вступає в реакцію з молекулою кисню. Під час відновлення Cu2+ до Cu+ за допомогою аскорбінової кислоти утворюється іон міді, здатний взаємодіяти з киснем з утворенням пероксидної сполуки. Далі гідроксильна група переноситься на молекулу дофаміну з утворенням норадреналіну.
Роль металів у регуляції активності ферментів. Іноді іони металів виступають у ролі регуляторних молекул. Так, іони Са2+ слугують активаторами ферменту протеїнкінази С, яка каталізує реакції фосфорилювання білків. Іони Са2+ також змінюють активність ряду кальційкальмодулінзалежних ферментів.
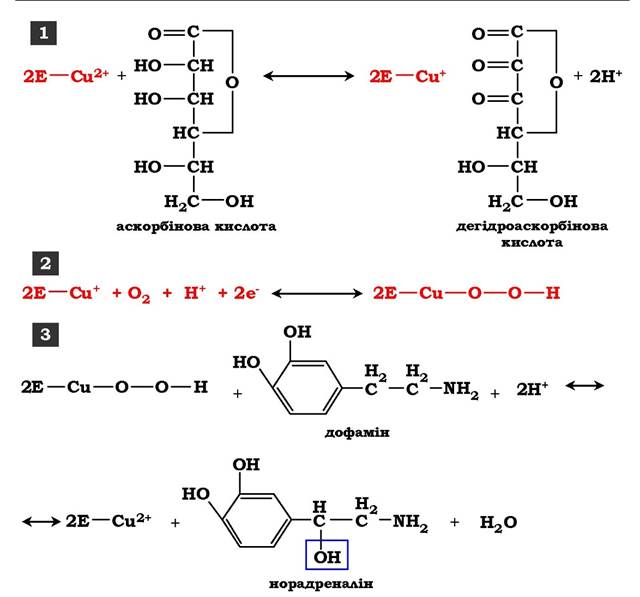
Рис. 7.6. Участь іону міді в активації молекули кисню під час функціонування дофамінгідроксилази:
1 - відновлення Cu2+, що входить до складу активного центру дофамінгідроксилази, до Cu+ за допомогою аскорбінової кислоти; 2 - взаємодія Cu+ з киснем з утворенням пероксидної сполуки; 3 - перенесення гідроксильної групи на молекулу дофаміну з утворенням норадреналіну