БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 9.НУКЛЕЇНОВІ КИСЛОТИ
9.4.Структура нуклеїнових кислот
9.4.2.Просторова організація структури ДНК
Дослідження нуклеотидного складу молекул ДНК із різних біологічних об'єктів показало, що незалежно від джерела походження (тваринні, рослинні, бактеріальні організми), усі ДНК мають певні кількісні співвідношення між вмістом пуринових і піримідинових нуклеотидів. Застосовуючи метод хроматографії, Е. Чаргафф і співробітники його лабораторії визначили нуклеотидний склад нуклеїнових кислот різного походження та дійшли висновку (1950 р.), що співвідношення в ДНК азотистих основ підпорядковуються універсальним закономірностям, які назвали правилами Чаргаффа:
✵ сума піримідинових нуклеотидів дорівнює сумі пуринових (Пур = Пір);
✵ кількість основ, які містять аміногрупи в положенні 4 піриміди- нового ядра та в положенні 6 пуринового (А + Ц), дорівнює кількості основ, які мають оксогрупу в тих самих положеннях (Г + Т);
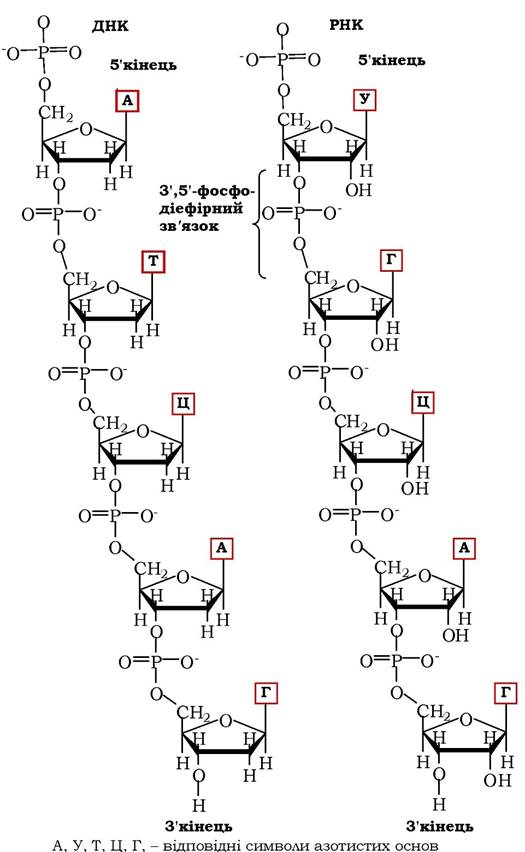
Рис. 9.5. Фрагменти полінуклеотидних ланцюгів ДНК і РНК
✵ молекулярні співвідношення Г + Ц / А + Т (коефіцієнт специфічності) видоспецифічні. Для ДНК вищих тварин і рослин та деяких мікроорганізмів це співвідношення менше 1 (у них так званий АТ-тип ДНК), у ДНК більшості мікроорганізмів, особливо бактерій і грибів, переважають ГЦ-пари (ГЦ-тип).
Ці емпіричні правила Чаргаффа стосуються тільки ДНК, і не властиві РНК.
Виявлені особливості кількісних взаємовідносин між азотистими основами вказують на те, що в ДНК повинна зберігатися досить чітка закономірність спарювання не пуринових і піримідинових основ узагалі, а відповідно - аденіну з тиміном і гуаніну з цитозином.
Використовуючи правила Чаргаффа, а також результати вивчення будови молекул ДНК методом рентгеноструктурного аналізу Дж. Уотсон і Ф. Крік у 1953 р. запропонували модель структури молекули ДНК у вигляді подвійної спіралі, яка має епохальне значення. Існує віршована версія книги Дж. Уотсона "Подвійна спіраль", написана Дж. Філдом (переклад Оксани Берник) про надзвичайне значення відкриття просторової структури ДНК.
Слухай пісню, як спіральний
Комплекс скручений, хіральний
Хтось відкрив і в світ пустив.
Цій конструкції
Після ретельної дедукції
Життя дали.
З усіх сторін: "О! Диво, люди!" —
Несамовито люд кричав.
А автор ручку взяв й писав
Чудову байку про спіральку:
Як кралечка із Кембриджа
Світ науковий ненароком
Із ніг на голову з прискоком
В прекрасну мить перевела.
Неначе сон прийшла вона.
Я вам всю правду розповім
Як ту структуру відкривали:
Загадку гена розгадали!
Біопрогнози і без містики
Краще будь-якої белетристики.
В думок великих буревії
Всі здогадки "прийшли до дії":
Пари основ миготіли в тумані
Всім перешкодам на зло.
Так воно все і було.
Легким помахом руки
Поєднати ці основи
Н-зв'язки допомогли.
Затьмарено всі експерименти
У хвалу звучать аплодисменти!
Як його не величати
Все ж сором хімії не знати
Та, прочитав він, так скажу,
Книгу Полінга одну.
Тих основ узяв моделі
Крутив їх наче в каруселі.
Клав їх так, і ставив сяк,
Та не слухались ніяк
Ні тимін, ні аденін,
Десь утік гуанозин.
Не покладав він рук щоденно,
А тут і лист із Пасадени.
Що тут коїться? О, Боже,
Полінга модель пригожа
Для атомів, так, додаткових,
Що в усіх посібниках зразкових
Як і писали - тільки прикінці.
В образі
(може, в нецензурній фразі)
про модель він відгук дав
але рук, повір, не склав!
Почав знову працювати
Основи сяк і так в'язати.
Що ж вам далі розказати?
Автор наш таутомери
(Те що треба) вибирає
Сама природа - помагає.
Ланцюжки два (чи спіралі)
Плюс іще такі деталі:
Полінга основи ззовні,
А фосфати посередині,
Навпаки всі факти вам наведені.
R-шляхом перевіряли,
Щоб на сто відсотків знали!
...Ось і про наше відкриття,
Догматичної природи,
Слухали усі народи.
Важко було: Села Розі
На рефлекси в гордій позі.
Книгу Брегг назвав скрижаллю Моріс? —
Гнівно і без жалю,
Наче чайник закипів
Далі можна і без слів.
Що за байка без моралі:
Ось пиши такі скрижалі,
Часу не гайнуй намарне —
Науковцем станеш гарним.
А тут і НОБЕЛІВСЬКА премія —
Побачить світ іще одного генія!
Згідно з моделлю Уотсона - Кріка, молекула ДНК складається з двох полінуклеотидних ланцюгів, що утворюють правообертальну спіраль, яка має загальну вісь. Азотисті основи спрямовані всередину спіралі, а їхні площини майже перпендикулярні її осі й паралельні одна одній.
Між основами за такого розміщення виникає міжплощинна взаємодія - стекінг (англ. stack up - розташування один над одним). При цьому забезпечуються не тільки вигідні вандер-ваальсові контакти між атомами, але й виникає додаткова стабілізація завдяки перекриванню π-орбіталей атомів контактуючих основ. Стабілізація подвійних спіралей ДНК здійснюється також за рахунок сприятливого гідрофобного ефекту, який виявляється в тому, що неполярні основи захищенні від безпосереднього контакту з водним (гідрофільним) оточенням. Навпаки, вуглеводно- фосфорний каркас з його полярними групами та зарядженими атомами експоновані, що також стабілізує структуру й забезпечує формування вторинної структури ДНК.
Два ланцюги молекули ДНК мають протилежну спрямованість - один із них орієнтований у 3' → 5' напрямку, а другий - у 5' → 3'. Подібна протилежна полярність двох ланцюгів у спіралі забезпечує правильну просторову взаємну орієнтацію азотистих основ.
Полінуклеотидні ланцюги в молекулі ДНК, які орієнтовані антипаралельно, по всій довжині зв'язані один з одним водневими зв'язками. Ці зв'язки між ланцюгами утворюються за рахунок специфічної взаємодії аденінового залишку одного ланцюга з ти- міновим залишком другого ланцюга (два водневих зв'язки) та гуанінового залишку одного ланцюга з цитозиновим залишком другого ланцюга (три водневі зв'язки). Нижче вказані місця водневих зв'язків між основами в нуклеїнових кислотах (рис. 9.6):
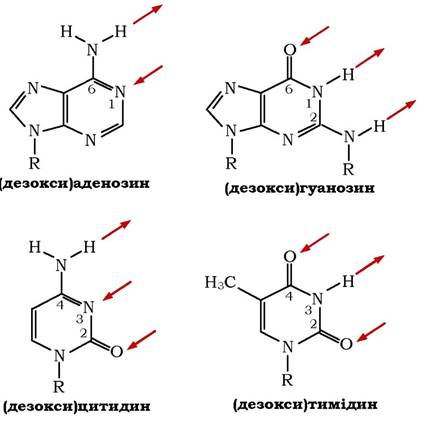
Рис. 9.6. Місця водневих зв'язків азотистих основ у нуклеїнових кислотах
Азотисті основи, які утворюють пари, комплементарні одна одній в тому відношенні, що між ними легше виникають водневі зв'язки, ніж при інших поєднаннях. Відбувається це тому, що в цих парах центри з підвищеною та пониженою електронною щільністю основ розміщенні оптимально один до одного.
У дволанцюговій молекулі ДНК стійкість конформації зумовлена обмеженим обертанням навколо фосфодіефірного зв'язку й забезпечується переважаючою антиконформацією глікозидних зв'язків і домінуючими таутомерними формами п'яти азотистих основ (див. рис. 9.1 і 9.2)
Спіраль ДНК регулярна: один виток спіралі (360°) складається з 10 нуклеотидних пар, ця структура повторюється з інтервалом у 3,4 нм, відстань між азотистими основами вздовж осі спіралі становить 0,33 нм, а діаметр - 2,37 нм. Кожна основа повернена відносно попередньої на 36°.
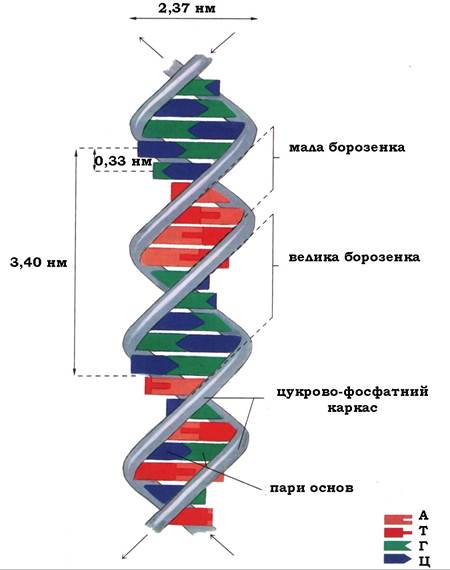
Рис. 9.7. Схема будови подвійної спіралі ДНК (В-форма)
Просторове взаєморозміщення ланцюгів зумовлює виникнення великої та малої борозенок, завширшки 2,2 нм і близько 1,2 нм відповідно. У ділянці великої борозенки ДНК асоційовані регуляторні білки, які можуть специфічно взаємодіяти з певними атомами азотистих основ і здійснювати контроль експресії генів, не порушуючи при цьому комплементарних взаємодій у структурі подвійної спіралі (рис. 9.7).
Подвійні структури в молекулах ДНК виникають не тільки при взаємодії двох комплементарних полідезоксирибонуклеотидних ланцюгів, але й у межах одного й того ж ланцюга. Це відбувається в тих випадках, коли в комплементарних ланцюгах присутні інвертовані послідовності (паліндроми, грец. palindrome - перевертень), тобто нуклеотидні послідовності, що читаються однаково у прямому та зворотному напрямках. Наприклад, послідовність
![]()
є паліндромом, оскільки при зчитуванні від 5' до 3' вона ідентична (ГААТТЦ) в обох ланцюгах. Особлива властивість таких обернених ділянок ДНК полягає в тому, що за певних станів ДНК вони здатні утворювати шпилькові структури простої або складнішої форми (рис. 9.8).
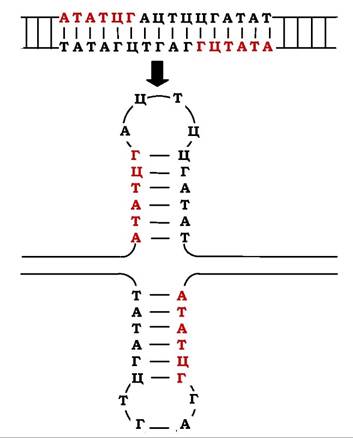
Рис. 9.8. Приклад інвертованих нуклеотидних послідовностей та їхня здатність формувати різні структури ДНК
Практично кожна ДНК містить паліндромні послідовності довжиною від декількох до багатьох тисяч пар нуклеотидів. Паліндромні структури характерні для тих ділянок молекули ДНК, де розташовані зони впізнавання структур ДНК ферментами та регуляторними білками.
Зазначені структурні особливості дволанцюгової молекули ДНК характерні для В-форми молекули ДНК, яка є домінуючою за фізіологічних умов (низька концентрація солі, висока ступінь гідратації). Однак при взаємодії нуклеїнової кислоти з різною кількістю молекул води, катіонами та іншими факторами ДНК набуває різних форм. На цей час ідентифіковані та описані такі форми ДНК: правообертальні А-, В-, С-, D-, Е-, Н-, L-, Р- і лівообертальна спіраль - Z-. Проте в природних біологічних системах спостерігалися тільки А-, В- і Z-форми ДНК.
Подібний поліформізм молекули ДНК пов'язаний з:
✵ кількістю пар основ, які припадають на один виток подвійної спіралі;
✵ відстанню між площинами пар основ і кутом, який вони утворюють із віссю спіралі;
✵ діаметром спіралі;
✵ направленістю (права, ліва) подвійної спіралі, яка залежить від послідовності нуклеотидів в ДНК, величини та напрямку її суперспіралізації, хімічної модифікації азотистих основ і концентрації хімічних речовин у розчині, перш за все іонів металів і поліамінів.
Деякі геометричні дані ізоформ ДНК наведено в табл. 9.3.
Таблиця 9.3
Характеристика деяких ізоформ ДНК
Форма ДНК |
Кількість пар основ на виток |
Діаметр спіралі, нм |
Відстань між площинами основ, нм |
А |
11 |
2,3 |
0,256 |
Праві спіралі В |
10 |
1,9 |
0,338 |
С |
9 |
1,9 |
0,320 |
Ліва спіраль Z |
12 |
1,8 |
0,371 |
Зміна конфігурації ДНК залежно від умов, в яких перебуває молекула, зумовлена тим, що валентні кути між основами й пентозами можуть змінюватися, і пентозофосфатний каркас є досить гнучким для того, щоб утворювались альтернативні форми подвійної спіралі ДНК. Слід відмітити, що деякі з цих форм переходять одна в одну в разі зміни будь-яких вказаних вище параметрів та дії певних факторів. У клітині ДНК зазвичай існує у В-формі, але окремі її ділянки можуть перебувати в А-, Z- або в іншій конформації. Важливо, що А-форма ДНК схожа зі структурою, характерною для дволанцюгової РНК, або для гібридних молекул ДНК-РНК. Для наочності наводимо схему кількох альтернативних форм (рис. 9.9).
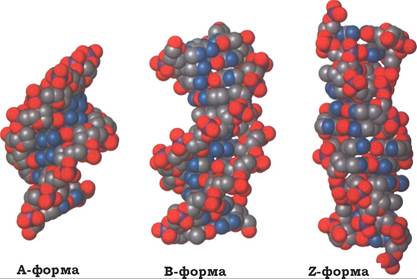
Рис. 9.9. Ізоформи ДНК
За певних умов ділянки ДНК, для яких характерним є чергування пуринових і піримідинових нуклеотидів (ГЦ або АЦ), набувають форми лівої спіралі (Z-форма). При цьому відстань між сусідніми парами основ збільшується до 0,371 нм (див. табл. 9.3), а кількість пар основ на один виток - до 12. Каркас молекули ДНК має зигзагоподібний вигляд унаслідок того, що переміжні нуклеотидні послідовності гуаніну й цитозину та залишки дезоксирибоз зазнають конформаційних змін, через що лінія, яка сполучає фосфатні групи, через кожні дві пари основ утворює злам.
До стабілізуючих Z-форму ДНК факторів можна віднести: 1) зв'язування Z-ДНК-специфічних білків; 2) метилюванння атомів вуглецю в 5-положенні деяких залишків дезоксицитидину; 3) наявність специфічних катіонів, таких як спермін і спермідин та ін. Припускають, що ДНК у Z-формі може брати участь у регуляції експресії генів, які близько розташовані, а також суттєво віддалених від Z-ділянки.
Лінійна ДНК у клітині має форму витягнутої молекули, упакованої в компактну структуру з утворенням спіралізованих і надспіралізованих форм. Це свідчить, що дволанцюгова молекула ДНК у просторі може піддаватись подальшій укладці в певну третинну структуру - надспіраль, або кільцеву форму. Бактеріальні плазміди, хромосоми деяких бактерій, більшість мітохон- дріальних і хлоропластних ДНК та ДНК-вмісник вірусів тварин представлені ковалентно замкненими кільцевими молекулами ДНК. Замкнуті кільця можуть існувати в релаксованому або надспіралізованому стані (рис. 9.10).
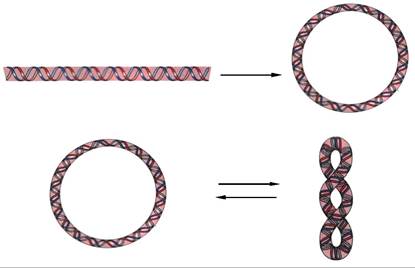
Рис. 9.10. Лінійна, релаксована циклічна і надспіралізована форми ДНК
При перетворенні релаксованої структури ДНК у надспіралізо- вану затрачується певна кількість енергії й молекула стає компактнішою, що дає можливість упакування молекули ДНК у малий об'єм. Надвитки в ДНК, утворені за рахунок спіралізації проти годинникової стрілки у напрямку, зворотному закручуванню правосторонньої подвійної спіралі ДНК, називаються негативними. При переході молекули ДНК до іншого типу надмолекулярної структури, енергія може знижуватися за рахунок утворення ділянок негативної спіралізації. Прикладом такого переходу є розділення ланцюгів ДНК під час матричних процесів - синтезу ДНК (реплікація) або синтезу РНК (транскрипція). Ферменти, які каталізують топологічні зміни молекули ДНК, називаються топоізомеразами.
Надспіральна конформація ДНК характерна для нуклеоїду прокаріотів і хромосом вищих організмів. Подібна третинна структура стабілізується за рахунок електростатичних сил - негативно заряджені фосфатні групи частково нейтралізуються позитивно зарядженими іонами металів і поліамінами або основними амінокислотними залишками білків. У результаті таких взаємодій відбувається конденсація ДНК зі зменшенням об'єму в тисячі разів. Наприклад, довжина молекули ДНК однієї з найменших хромосом людини становить близько 3 см, а сумарна довжина всієї ДНК однієї клітини складає близько 2 м. При цьому вона упакована в ядрі з діаметром близько 5 мкм.
Кожна молекула ДНК упакована в окрему хромосому, до складу якої входить аморфне нуклеопротеїнове утворення - хроматин, стан якого змінюється залежно від клітинного циклу. У фазі спокою хроматин рівномірно розподілений по всьому об'єму ядра. У фазі поділу клітини хроматин утворює компактні частки - хромосоми. Морфологічно розрізняють еухроматин і гетерохроматин, який більш конденсований, ніж еухроматин. Останній відповідає ділянкам хромосом з активною транскрипцією.
Хроматин - надмолекулярна структура, що складається з ДНК, гістонів та інших ядерних білків (негістонових білків). Приблизно 2/3 маси хроматину становлять білки, 1/3 - ДНК; хроматин містить також РНК (до 10 %), незначний відсоток ліпідів та іони деяких металів (Mg2+, Ca2+, Fe2+). Терміном "гістони" позначають декілька груп споріднених основних білків із молекулярною масою 11-21 кД. Характерною особливістю гістонів є високий вміст лізину і/ або аргініну. Завдяки позитивному заряду гістони утворюють іонні зв'язки з негативно зарядженими фосфатними групами, які розташовані на зовнішній поверхні подвійної спіралі ДНК.
Існує п'ять основних типів гістонів: Н1, Н2А, Н2В, Н3 та Н4. По дві молекули кожного з гістонів Н2А, Н2В, Н3 і Н4 становлять октамер (нуклеосомний кор) (рис. 9.11, А), оповитий сегментом ДНК довжиною 146 пар нуклеотидів (1,75 оберта). Такий комплекс гістонових білків з ДНК є основною структурною одиницею хроматину й називається нуклеосом.ою. У складанні нуклеосом бере участь ядерний білок аніонного характеру - нуклеоплазмін, здатний оборотно сполучатися з гістоновим октамером і таким чином блокувати здатність гістонів до неспецифічної взаємодії з негативно зарядженими структурами, такими як ДНК. Нуклеоплазмін проявляє вибірковість до певних ділянок ДНК і, мабуть, створює в ядрі специфічне іонне оточення, яке сприяє взаємодії гістонів з ДНК і складанню нуклеосом.
Ділянка ДНК, розташована між сусідніми нуклеосомними частками, називається лінкерною (сполучною) ДНК, і в середньому складається з 60 пар нуклеотидів. Молекула гістону Н1 не входить до складу нуклеосомного кору й не бере участі в процесі намотування ДНК на гістоновий октамер. Гістон Н1 контактує з ДНК у тих місцях, де подвійна спіраль входить і виходить із нуклеосомного кору, тобто гістон Н1 "зшиває" ДНК у місцях, де вона починає і припиняє обвиватися на нуклеосомний кор (рис. 9.11, Б).
Подальше укладання нуклеосом у ядрі залежить, мабуть, від взаємодії гістонів Н1 з ділянками дволанцюгової ДНК, що сполучає нуклеосоми. Топологію цієї взаємодії, яка приводить до утворення міжнуклеосомних з'єднань, вивчено недостатньо. Але за допомогою електронної мікроскопії в інтерфазному ядрі було виявлено соленоїдоподібну структуру діаметром 30 нм, або, як її ще називають, 30 нм-фібрила.
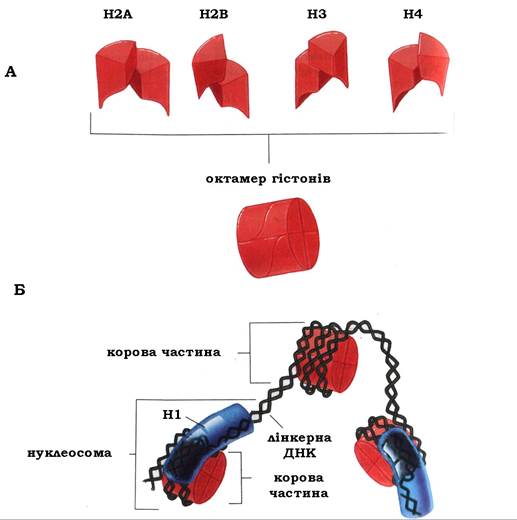
Рис. 9.11. Модель структури нуклеосомного кору (А) і нуклеосом (Б)
Припускають, що дископодібні нуклеосоми діаметром 10 нм і висотою 5 нм у цій упаковці дотикаються одна до одної краями й орієнтуються своїми плоскими поверхнями вздовж осі фібрили. Фібрили, імовірно, також спіралізуються з кроком 6 нуклеосом на один виток спіралі. У результаті утворюється соленоїд діаметром 30 нм (рис. 9.12).
Коли хроматин конденсується з утворенням метафазної хромосоми, соленоїдні структури утворюють петлеподібні домени, діаметром 300 нм, зв'язані з білковим каркасом (ядерний каркас). Ядерний каркас являє собою хромосомні структурні білки, на яких відбувається остаточна конденсація хроматину.

Рис. 9.12. Модель 30 нм-хроматинової фібрили наднуклеосомного рівня укладання ДНК
Наступна компактизація ДНК приводить до утворення додаткових надструктур. Приблизно 20 петель утворюють так звані мінідиски, які, у свою чергу, укладаються у стосик (700 нм) (рис. 9.13).
Наслідком такого упакування ДНК є утворення конденсованого хроматину, що є характерним для еукаріотичної метафазної хромосоми і приводить майже до 8000-10000-кратного укорочення молекули ДНК.
Деякі амінокислотні залишки (аргінін, лізин) і кінцеві аміногрупи гістонів у складі нуклеосом здатні хімічно модифікуватися: фосфорилюватися, метилюватися, ацетилюватися або взаємодіяти з білком убіквітином тощо. Такі ковалентні модифікації можуть викликати зміни заряду й конформації гістонів, що, у свою чергу, впливає на взаємодію гістонів між собою та з ДНК. Хімічні модифікації роблять можливими конфірмаційні перебудови хроматину, що є важливим для контролю за генною експресією.
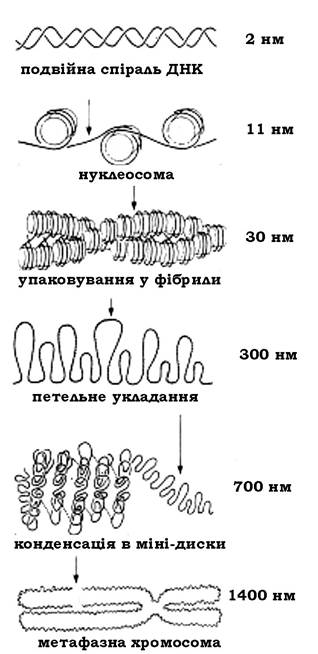
Рис. 9.13. Схема компактизації ДНК у хромосомах
Негістонові білки хроматину. В ядрі еукаріотичних клітин присутні різноманітні ДНК-зв’язуючі негістонові білки. До них належать:
✵ білки, які виконують структурну функцію, беруть участь в утворенні наднуклеосомних рівнів укладання хромосом;
✵ численна група ферментів, що забезпечують процеси реплікації, транскрипції, репарації, хімічної модифікації складових компонентів хроматину;
✵ найрізноманітніша за складом група регуляторних білків, які контролюють активність указаних вище ферментів, а також доступність тих чи інших ділянок ДНК для цих ферментів. Як приклад можна навести родину сайт-специфічних білків типу "цинкові пальці», лейцинові "застібки", гомодимери. Вони мають особливу структуру та належать до так званих факторів транскрипції, які зв'язуються з регуляторними областями генів і таким чином впливають на експресію. До групи структурно-регуляторних білків, постійно асоційованих із хроматином, відносять білки високої рухомості - HMG-білки (від англ. high mobility gel proteins). Ці білки характеризуються відносно малими розмірами (до 30 кД) і високим вмістом заряджених амінокислотних залишків. Вони виявляють спорідненість і зв'язуються з тими нуклеосомами, які містяться в транскрипційно-активній частині хроматину.