БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 10. МЕТАБОЛІЗМ НУКЛЕЇНОВИХ КИСЛОТ
10.7.Біосинтез РНК (транскрипція)
10.7.3.Процесинг РНК-транскриптів
Утворені під час транскрипції первинні транскрипти різних видів РНК надалі формуються в дозрілі молекули, які виконують відповідні функції. Післятранскрипційний процес перетворення попередників РНК у функціонально-активні молекули називається процесингом. Він передбачає розщеплення довгих первинних транскриптів, видалення одних і приєднання до 3'-,5'-кінців інших нуклеотидів, хімічну модифікацію нуклеотидів.
Процесинг молекул РНК відбувається у прокаріотичних і в еукаріотичних організмів, але особливо складні перетворення попередників клітинних РНК у ядрах еукаріотів. У багатьох генах еукаріотичних організмів міститься два різновиди нуклеотидних послідовностей - екзони та інтрони. Екзони - це ділянки структурних генів, в яких закодована нуклеотидна послідовність функціонально активних (дозрілих) РНК. Між екзонами розташовані протяжні ділянки ДНК, які не несуть генетичної інформації безпосередньо в амінокислотну послідовність. Такі послідовності називають інтронами. Переважна більшість первинних транс- криптів структурних генів включають ділянки як екзонів, так і інтронів. Під час дозрівання РНК-попередників фрагменти, які відповідають інтронам видаляються, а структурні частини (екзони) ковалентно сполучаються в безперервний ланцюг. Даний процес називається сплайсингом (англ. splice - сполучати, зрощувати). Для його здійснення необхідні високоспецифічні ендонуклеази, які з високою точністю (до одного нуклеотиду) каталізують розрив фосфодіефірного зв'язку на межі екзон-нітрон, та спеціальні лігази, що сполучають екзони.
Дотримання точності цього процесу є вельми важливим, оскільки в протилежному випадку помилка навіть на один нуклеотид може викликати зміщення "рамки" трансляції, що приводить, наприклад, до утворення термінуючих триплетів і до зупинки синтезу білка.
Проходження більшістю РНК-транскриптів через стадію процесингу є одним із регуляторних механізмів, тобто дає змогу регулювати експресію генів на посттранскрипційному рівні.
Процесинг попередників мРНК. У прокаріотичних організмів первинні транскрипти мРНК-кодуючих генів починають використовуватися як матриці білкового синтезу ще до завершення транскрипції, без проходження ними хімічних модифікацій або процесингу.
Молекули еукаріотичних мРНК зазвичай утворюються з більших за розміром молекул-попередників, які отримали назву гетерогенної ядерної РНК (гяРНК). Для утворення дозрілої мРНК ці молекули піддаються ковалентним модифікаціям по 5'- та 3'-кінцях і сплайсингу.
Модифікація 5'-кінця попередника мРНК починається ще на стадії елонгації, коли довжина первинного транскрипту сягає приблизно 30 нуклеотидних залишків. У цей час відбувається кепування 5'-кінця попередника. Реакцію каталізує фермент гуанілтрансфераза, який гідролізує макроергічний зв'язок у молекулі ГТФ і приєднує нуклеозидмонофосфатний залишок 5'-фосфатною групою до 5'-кінця синтезованого фрагмента РНК з утворенням не характерного для нуклеїнових кислот 5',5'-фосфо- діефірного зв'язку між нуклеотидами. Завершується формування кеп-структури метилюванням залишку гуаніну в складі ГТФ з утворенням N 7 - метилгуанозину. У кеп-структурі піддаються також метилюванню рибозні залишки нуклеотидів.
Модифікований 5'-кінець забезпечує захист мРНК від дії 5'-екзо- нуклеаз і фосфатаз, транспорт мРНК у цитоплазму, забезпечує ініціацію синтезу білка; останнє особливо важливо, оскільки ініціюючі триплети (АУГ, ГУГ) розпізнаються рибосомою тільки за наявності кеп-структури.
Модифікація З'-кінця попередника мРНК здійснюється по- лі(А)-полімеразою, яка понуклеотидно нарощує поліаденіловий фрагмент (полі (А)-"хвіст"), що складається зі 100-200 залишків аденілової кислоти. Поліаденілювання відбувається за наявності в РНК-транскрипті специфічної нуклеотидної послідовності - ААУААА (полі-А-сигнал). Наявність подібного сигналу призводить до активації поліаденілатполімерази, яка спочатку каталізує розщеплення З'-фосфодіефірного зв'язку в ланцюзі РНК, а потім до З'-кінця в точці розриву фермент нарощує полі(А)-фрагмент у кількості до 200 нуклеотидів.
Наявність полі (А)-послідовності на З'-кінці полегшує вихід мРНК із ядра й уповільнює її гідроліз у цитоплазмі, а також сприяє підтриманню внутрішньоклітинної стабільності цієї молекули.
Процес сплайсингу. Первинний транскрипт мРНК, що комплементарний гену, містить як екзони, так і інтрони. Послідовності інтронів вирізаються з первинного транскрипту, а кінці екзонів ковалентно сполучаються один з одним з утворенням дозрілих молекул (рис. 10.24). Порядок видалення інтронів необов'язково відповідає послідовності розташування їх у гені. Кількість інтронів може сильно варіювати - від одного до декількох десятків, і вони іноді становлять більшу частину гена. Сумарна довжина нуклеотидів інтронів може в багато разів перевищувати довжину екзонів.
Видалення інтронів здійснюється спеціальними ендонуклеазами й за участю малих ядерних рибонуклеопротеїнів (мяРНП), які складаються з мяРНК, ланцюг якої зв'язаний з білковим каркасом, що складається з декількох протомерів. Роль мяРНП полягає в тому, що вони за принципом комплементарності вибірково асоціюються з 5'- та 3'-послідовностями сайтів сплайсингу первинного транскрипту й каталізують реакцію розщеплення 3', 5'-фосфодіефірного зв'язку на кордоні екзона з інтроном. Послідовності інтронів видаляються, а екзони сполучаються ферментом РНК-лігазою.
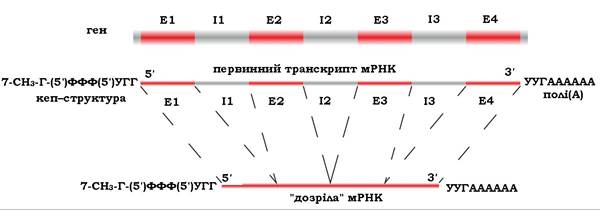
Рис. 10.24. Схема сплайсингу мРНК:
Е1, Е2, Е3, Е4 - екзони; І1, І2, ІЗ — інтрони
У деяких випадках відбувається альтернативний сплайсинг і поліаденілювання одного й того ж транскрипту. Для таких генів характерним є те, що екзон одного варіанта сплайсингу може виявитися інтроном і тому такі молекули мРНК будуть різнитися набором екзонів. Це приводить до утворення різних мРНК і, відповідно, різних білків з одного первинного транскрипту.
Процес сплайсингу відбувається в ядрі, у цитоплазму переноситься вже дозріла мРНК, яка знаходиться в комплексі з білками. Інтронні фрагменти розщеплюються нуклеазами до окремих нуклеотидів, які знову використовуються для синтезу РНК у ядрі.
Процесинг попередників рРНК. Дозрілі молекули рРНК прокаріотичних організмів утворюються зі спільного попередника з коефіцієнтом седиментації 30S, який містить майбутні дозрілі 16S, 23S, 5S рРНК і тРНК (рис. 10.25).
Загалом РНК-транскрипт піддається дії нуклеаз, специфічність дії яких значною мірою зумовлена вторинною структурою попередника. Початкове розщеплення транскрипту здійснюється ендонуклеазою ІД яка атакує дволанцюгові ділянки РНК, утворені завдяки внутрішньомолекулярному спарюванню. У результаті здійснюється одночасне дозрівання 5'- та 3'-кінців рРНК. Услід за РНКазою ІІІ, певно після взаємодії РНК- транскрипту з рибосомними білками, у процесингу беруть участь М-ендонуклеази (англ. maturation - дозрівання) М5, М16, М23. Дія всіх ендонуклеаз супроводжується утворенням 5'-фосфатних і 3'-гідроксильних кінців.
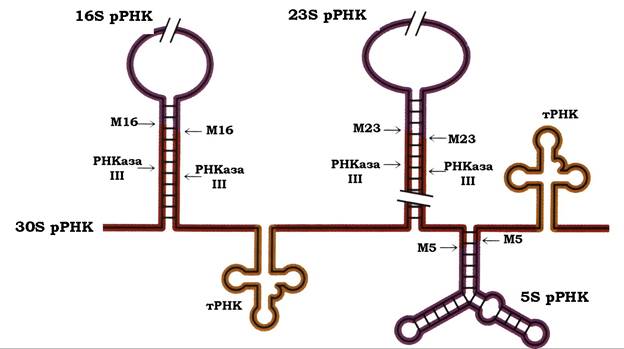
Рис. 10.25. Процесинг попередника рРНК E. coli: М — ендонуклеази
Описана вище багатоступеневість процесингу рРНК може бути пояснена можливістю регулювати на кожній стадії швидкість утворення зрілих рРНК. Крім того, суттєвим є ще й те, що локалізація в одному первинному транскрипті майбутніх 23S і 16S рРНК, які входять до складу малої та великої субодиниць рибосом, автоматично забезпечує утворення їх у рівних молярних співвідношеннях.
У клітинах еукаріотичних організмів первинний транскрипт рРНК (45S) містить послідовності відразу трьох дозрілих рРНК - 18S, 5,8S і 28S. Ці послідовності розділені спейсерами, але не містять інтронів. Спейсери (англ. spacer - роздільник) - це міжгенні нетранскрибуючі нуклеотидні послідовності, які розділяють повторювальні транскрибуючі елементи генного кластера (рис. 10.26). У спейсерах відсутні модифіковані нуклеотиди, що містяться в дозрілих рРНК.
Загальний попередник рРНК піддається процесингу, під час якого метилюються деякі рибозні залишки та утворюються 18S рРНК, яка входить до складу малої субодиниці рибосом, а також 28S і 5,8S рРНК, які локалізуються у великій субодиниці рибосом. Решта транскрипту руйнується в ядрі.
Усі три рРНК утворюються в рівних кількостях, оскільки вони походять з одного й того ж первинного транскрипту.
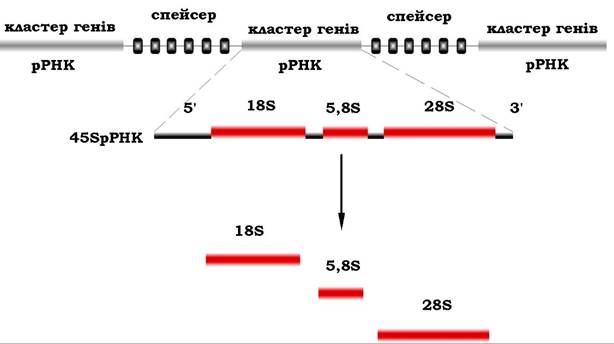
Рис. 10.26. Процесинг попередника рРНК еукаріотів
До складу великої субодиниці рибосом входить ще 5S рРНК, яка транскрибується окремо від первинного транскрипту 45S рРНК.
Процесинг попередників тРНК. Функціонально активні молекули тРНК про- та еукаріотів утворюються з більш довгого попередника, який піддається розщепленню та модифікації з включенням мінорних основ.
Незважаючи на те, що деякі кодуючі тРНК гени прокаріотів містяться всередині транскрипційних одиниць рРНК і експресу- ються спільно з генами рРНК (див. вище процесинг попередників рРНК та рис. 10.25), основна частина тРНК-генів представлена поодинокими генами або вони об'єднані в кластери. Однак незалежно від цього, 5'-кінці всіх тРНК-транскриптів утворюються за участю ендонуклеази - РНКази Р (від англ. precursors - попередник), яка впізнає певну вторинну ("лист конюшини») або третинну (Г-подібну) структури молекули тРНК (рис. 10.27, А).
Характерною особливістю РНКази Р є те, що вона складається з білка та РНК, яка містить від 100 (людина) до 375 (Е. coli) нуклеотидів. Як з'ясувалося, саме РНК каталізує дозрівання попередників тРНК, а білкова складова ферменту тільки підсилює каталітичну активність РНК шляхом утримання структури РНК в максимально активній конфігурації. Дія РНК у складі РНКази Р проявляється як справжній каталізатор, властивості якого визначаються нативною структурою РНК. Такі каталітичні РНК мають назву рибозими.
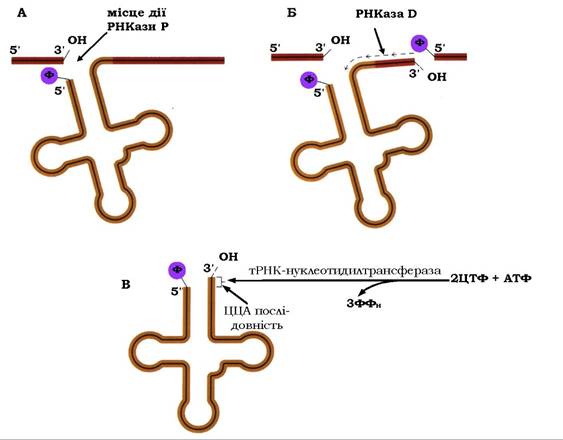
Рис. 10.27. Процесинг попередника тРНК
Процесинг 3'-кінця тРНК-транскрипту здійснюється за допомогою РНКази D (від англ. digestion - перетравлення), яка являє собою екзо-3'-нуклеазу. Це непроцесивний фермент, при функціонуванні якого комплекс з субстратом дисоціює після кожного акту відщеплення нуклеотиду (рис. 10.27, Б). Ферментативне відокремлення нуклеотидів із 3'-кінця триває до моменту, коли нуклеаза досягає тринуклеотидної послідовності ЦЦА. При дозріванні деяких видів тРНК під впливом екзонуклеази утворюється кінець, який править приманкою, до якої тРНК-нуклеотидилтрансфераза послідовно приєднує ЦЦА до 3'-кінця (рис. 10.27, В).
Особливість процесингу тРНК в еукаріотичних організмів зумовлена тим, що ядерні попередники містять одиничний інтрон довжиною 10-40 нуклеотидів, розташований відразу за антико- доном (від 5'-кінця тРНК) (рис. 10.28, А). Тому процесинг первинних транскриптів багатьох тРНК-молекул потребує видалення інтрона і точного сплайсингу. Нуклеолітичний процесинг попередників тРНК, можливо, спрямовується не власне нуклеотидною послідовністю, а особливою тривимірною структурою, яку спроможні формувати молекули тРНК.
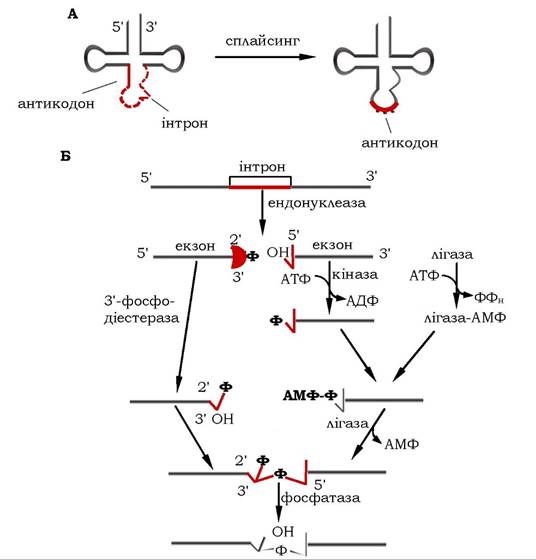
Рис. 10.28. Сплайсинг попередника тРНК в еукаріотів:
А - загальна схема, Б - механізм реакції
Механізм реакцій сплайсингу попередника тРНК за участю ферментів зображено на рис. 10.28, Б. Спочатку ендонуклеаза розщеплює РНК у двох сайтах сплайсингу, утворюючи на кінцях екзонів 5'-ОН групи та 2'-3'-циклофосфат. Наступна кіназна реакція за участю АТФ призводить до фосфорилювання 5'-кінця, а 3'-кінець піддається дії 3'-фосфодіестерази, унаслідок чого циклічна фосфатна група розкривається й утворюється 2'-фосфатна та 3'-ОН-групи.
З'єднання двох кінців екзонів відбувається за участю РНК- лігази, яка каталізує утворення фосфодіефірного зв'язку. Для цього вона спочатку активується шляхом аденілювання з утворенням аденільованого білка, а потім відбувається перенесення АМФ із ферменту на 5'-фосфатну групу екзона з утворенням 5',5'- фосфатного зв'язку. Далі 3'-ОН-група другого екзона витискує АМФ з 5'-кінця, а за впливу фосфатази відбувається дефосфорилювання в 2'-положенні.
Важливим етапом дозрівання попередника тРНК є різноманітні реакції ковалентних модифікацій. Наприклад, певні залишки уридину піддаються відновленню з утворенням дигідроуридину, ізомеризації з утворенням псевдоуридину, метилюванню з утворенням метилуридину; деякі залишки аденозину дезамінуються, перетворюючись в інозин, який потім може піддаватися метилюванню - утворюється метилінозин.
Весь спектр перетворень приводить до утворення декількох десятків видів тРНК - по 1-3 і більше для кожної із 20 амінокислот.