БІОХІМІЯ - Підручник - Остапченко Л. І. - 2012
Розділ 15. ІНТЕГРАЦІЯ МЕТАБОЛІЧНИХ ШЛЯХІВ. ГОРМОНИ
15.6.Гормони аденогіпофіза
Гормони аденогіпофіза класифікують за будовою: 1) прості білки, 2) глікопротеїни, 3) похідні ПОМК (проопіомеланокортину).
Соматотропний гормон (СТГ) і пролактин (ПЛ) є простими білками, складаються з одного поліпептидного ланцюга й характеризуються значною подібністю первинної структури.
СТГ синтезується в соматотропних клітинах гіпофіза й на відміну від інших тропних гормонів не має своєї ефекторної ендокринної залози. Основною функцією гормону є забезпечення лінійного росту та анаболічна дія. СТГ людини містить 191 амінокислотний залишок і має два дисульфідні зв'язки. Рівень секреції СТГ залежить від співвідношення концентрацій соматоліберину й соматистатину. Потрапляючи у кров, СТГ взаємодіє з СТГ-зв'язувальним білком, який є гомологічним зовнішньому домену рецептора СТГ на мембрані. Рістстимулюючі ефекти СТГ опосередковані інсуліноподібними факторами росту (ІФР), котрі синтезуються переважно в печінці під його впливом. ІФР-1 та ІФР-ІІ - це близькі за будовою одноланцюгові білки, подібні до проінсуліну, які знаходяться в сироватці переважно у вигляді комплексів з ІФР-зв'язувальними білками. Рецептори ІФР-1 мають тирозинкіназну активність. Обидва ІФР беруть участь у розвитку плоду, у постембріональний період основну роль у регуляції росту відіграє ІФР-І, який стимулює проліферацію клітин усіх тканин, і в першу чергу - хрящової та кісткової. Крім того, гормон збільшує синтез білка, гальмує його розщеплення. Має також ліполітичну дію для забезпечення тканин енергією при голодуванні та натще, знижує чутливість тканин до інсуліну, що сприяє розвитку інсулінорезистентності. Як СТГ, так і ІФР діють на гіпоталамус і аденогіпофіз за принципом зворотного зв'язку. Крім соматостатину гальмують секрецію гормону росту прогестерон, глюкокортикоїди, ожиріння.
Гшерсекреиія СТГ у дорослих викликає акромегалію, гіперсекреція у дітей зазвичай призводить до гігантизму, а гіперсекреція на пізніх стадіях статевого розвитку - до високорослості. Майже в усіх хворих на акромегалію виявляють СТГ-секретуючі аденоми гіпофіза. У більшості випадків причиною трансформації соматотропних клітин є мутації гена білка G80. Мутантний білок безперервно стимулює аденілатциклазу, що приводить до посилення проліферації соматотропних клітин і збільшення продукції СТГ. Абсолютний або відносний дефіцит СТГ приводить до зниження продукції ІФР-І і є однією з найпоширеніших причин затримки росту в дітей. Крім того, можливі затримки росту, зумовлені резистентністю до СТГ, викликаної дефектами гена рецептора.
Пролактин містить 198 амінокислотних залишків і синтезується в лактотропних клітинах. Головна мішень - молочні залози. Пролактин стимулює ріст молочних залоз під час вагітності й лактацію після пологів. У цьому беруть участь також прогестерон, кортизол і деякі інші гормони. Секреція пролактину постійно гальмується гіпоталамічним дофаміном і підсилюється під впливом естрогенів, тироліберину та нервових імпульсів від сосків.
Родина глікопротеїдних гормонів включає фолікулостимулюючий гормон (ФСГ), лютеїнізуючий гормон (ЛГ) і тиреотропний гормон (ТТГ), які складаються з 2 субодиниць: α-субодиниці гормонів ідентичні й містять по 92 амінокислотні залишки, β-субодиниці мають різну структуру та молекулярну масу, яка залежить в основному від кількості вуглеводних залишків. Обидві субодиниці глікозильовані.
ФСГ і ЛГ синтезуються в гонадотропних клітинах, містять по 115 амінокислотних залишків у β-ланцюзі й регулюють синтез та секрецію статевих гормонів і гаметогенез. ФСГ у яєчниках стимулює секрецію естрогенів, ріст і дозрівання фолікулів. ЛГ визиває овуляцію й утворення жовтого тіла, синтез і секрецію прогестерону в яєчниках. ЛГ стимулює клітини Лейдіга та синтез тестостерону, а ФСГ стимулює клітини Сертолі в яєчках, сперматогенез і синтез інгібіну. Продукція ЛГ і ФСГ регулюється гонадоліберином, а також статевими гормонами шляхом зворотного зв'язку - позитивного та негативного.
ТТГ синтезується в тиреотропних клітинах, його основна функція пов'язана зі стимуляцією синтезу тиреоїдних гормонів у щитоподібній залозі. Зв'язуючись із рецепторами на мембранах тиреоїдних клітин, ТТГ через активацію аденілатциклази стимулює надходження неорганічного йоду в щитоподібну залозу, синтез тиреоглобуліну, окиснення йоду та йодування тирозинів у тиреоглобуліні. ТТГ також стимулює ріст залози та її кровопостачання. Синтез і секреція ТТГ контролюється тиреоліберином. На секрецію ТТГ стимулюючий вплив має стрес і підвищена температура. Регуляція синтезу та секреції ТТГ здійснюється за механізмом негативного зворотного зв'язку.
Родина похідних проопіомеланокортину (ПОМК) - це гормони, які синтезуються з однієї молекули-попередника. (рис. 15.8) Молекула ПОМК містить 256 амінокислотних залишків. Спочатку з N-кінця відщеплюється сигнальний пептид (26 амінокислотних залишків), а потім у результаті протеолізу утворюються кортикотропін або адренокортикотропний гормон (АКТГ), а- і β-меланоцитостимулюючі гормони (МСГ), β- та у-ліпотропіни (ЛПГ), ендорфіни.
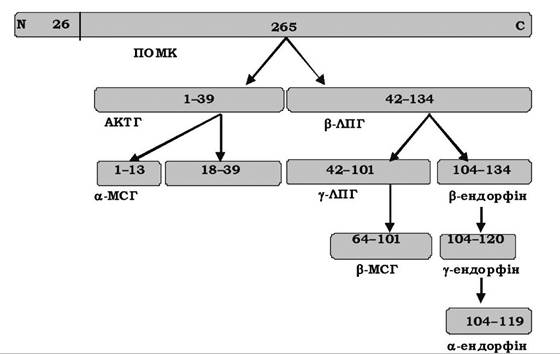
Рис. 15.8. Пептидні гормони, що утворюються з ПОМК
АКТГ складається з 39 амінокислотних залишків. Він стимулює функцію кори надниркових залоз, її ріст і забезпечення кров'ю, а також синтез гормонів у корі надниркових залоз, головним чином глюкокортикоїдів. Регуляція секреції АКТГ здійснюється стимулюючою дією кортиколіберину й негативним зворотним зв'язком з боку кортизолу. Кортизол також гальмує секрецію кортиколіберину.
МСГ стимулюють синтез меланіну у ссавців. Молекула β-ліпотропіну містить 91 амінокислотний залишок і виконує жиромобілізуючу дію. Шляхом специфічного протеолізу β-ліпотропіну утворюється ряд біологічно активних сполук опіатоподібної дії: метенкефалін, α-, γ-, δ- і β-ендорфіни. Найефективнішим анальгетиком є β-ендорфін.
Зниження секреції гормонів аденогіпофіза приводить до системних захворювань. Дефіцит ТТГ є причиною вторинного гіпотиреозу, дефіцит ЛГ і ФСГ приводить до вторинного гіпогонадизму, дефіцит АКТГ - до вторинної надниркової недостатності та гіпопігментації.